Exposing fallacies in ecological marine research in northern
New Zealand By Dr J Floor Anthoni (2003)
www.seafriends.org.nz/issues/cons/science.htm
Science is all about debate and discussion
of ideas. This section is highly critical of some marine ecological research
carried out in northern New Zealand waters, particularly the urchin barrens
hypothesis which has been wielded as a weapon in the marine reserves debate.
It is even taught at schools! Although this section intends to rebutt some
scientific findings, it is also valuable as study material to further one's
understanding of the ecology of the rocky shore in northern New Zealand
and what has gone wrong in marine ecological studies. Please note that
the principles involved, also apply to other places worldwide.
An introduction to the problems and why this section was created. An
explanation of top-down control and the ecologyy of urchin barrens. The
myth of unproductivity of barrens and the productivity of kelp.
Our own research into the extent of the kelpbed death of 1993, followed
by observations of its recovery gives us the strength to contest the findings
and conclusions of scientific research done on this subject.
Our version of the urchin barrens story is based on continual observation
of the marine reserve and nearby areas starting in 1976, but intensified
since establishing Seafriends in 1990, when we attained a boat. It allowed
us to study the kelpbed death in 1992/1993. We observed how the area recovered,
how urchins died and how isolated reefs did not recover until 1998. We
followed these observations up till today.
Protection of exploited fish in temperate regions: high density
and biomass of snapper Pagrus auratus (Sparidae) in northern New Zealand
marine reserves. Trevor J Willis, Russel B Millar, Russ C Babcock.
October
1997 to April 1999.
From time to time we will report changes to the environment as they
occur and any updates to this chapter.
related chapters on this web site
Storm
barrens of NZ: urchin barren zones are created by storms rather
than by urchins. Most storm barrens are not populated by urchins.
The snapper-urchin-kelp
myth: an explanation of the snapper 'trophic cascade' for the general
public. Your starting point.
The Dark Decay Assay:
a new plankton tool overturns our knowledge of the sea while quantifying
degradation and eutrophication. Important reading.
Survey93:
a
survey of the inner and outer Hauraki Gulf after the mass kelp die-off
of 1993. Floor Anthoni.
Bloom92:
The
algal bloom and climate anomalies of 1992. Dr Bill Ballantine.
Science needs skeptics:
why skeptics are an integral and necessary part of science
Science, technology
and human nature: the three culprits that caused all problems.
Why do we think they can save us?
The Goat Island marine
reserve: a virtual visit with history, ecology, biogeography, dive
spots, monitoring and more.
Introduction
to marine habitats: what it is like to live in the sea.
The ecology of Niue:
cyclones create very deep barrens on one side of Niue Island and many grazers
maintain these.
Disclaimer
When I started to doubt what
scientists were saying, already dating back to 1986, I could never have
foreseen that one day it would be necessary to dissect their studies in
order to expose glaring gaps and mistakes. I believed in the scientific
process of rigorous debate, peer review and criticism. I believed that
even their mistakes would eventually come to light as history has proved.
However, now that some blatant myths have been used as arguments for having
more marine reserves - worse still, they are being taught at school - the
time has come to expose these fallacies. We owe it to our children.
Floor Anthoni, June 2003.
When the search for truth is confused with
political advocacy, the pursuit of knowledge is reduced to
the quest for power. - Alston Chase
For comments, suggestions and improvements, please e-mail
the author, Dr Floor Anthoni.
For best printed results, read instructions
for printing.
-- seafriends home -- conservation
index -- Rev 20030731,20031211,20050803,20051219,20060926,20070315,20090521,
Introduction Marine research is difficult and arduous because of limited access
to the sea. As a result, the sea and its environments are some of the least
understood on this planet. Whereas ecological research has been hampered
by not being able to take the experiment into the laboratory, the sea is
worse still because it can also easily destroy experiments.
The following summary describes some of these concerns
The sea is hostile and difficult of access. Waves and weather are always
in control. We can be underwater only for one to a few hours each day.
Compare this with tramping the forests, and it is clear that we have very
little opportunity to become marine naturalists, but in the warm clear
tropical seas this gets better. Remember how much knowledge was gathered
by ecologists who were also keen naturalists?
As a result, the marine ecology is largely unknown. The plankton ecosystems
are also largely unknown, leaving wide gaps for new discovery as our revolutionary
Dark
Decay Assay method shows (2005).
With their limited general knowledge, scientists do 'controlled' ecological
experiments from which they derive far-reaching conclusions, which often
leads to nonsense when viewed from a wider perspective.
It is quite worrisome that the scientific null-hypothesis, in other words
proving that nothing else could have caused the effects, is no longer taken
serious. It is equally worrisome that experiments are not replicated before
publication.
Marine scientists do not have their own boats and are not free to dive
any place any time. They must account for every hour to some budget. They
have hardly enough time in the water to finish their own experiments. As
a result, very few experienced marine naturalists are found among marine
scientists.
Marine naturalists must be confident in diving, which takes the best of
ten years experience. They must learn to see, remember and understand the
functions of myriad creatures. After about 20 years one can begin to see
how it all works together and how it degrades. To recognise degradation
requires even more skills because what is no longer there, cannot be seen.
Marine research done in the lab is of high quality. But studying the ecology
cannot be done there. Controlled experiments are seldom possible. The scientific
method as defined by Francis Bacon fails there. Read Science,
technology & human nature. A different approach is needed but not
done (yet). FAQ.
Scientists cannot react to sudden events, even when these change the course
of their studies. First a budget is needed, and someone must be found to
pay for it. Time must be accounted for. As a result, many significant events
have not been studied and neither have these been mentioned, even when
it affected published results. Degradation has not been observed, measured
and published even though it has been around for nearly a century. Degradation
of coral reefs is often ignored. Degradation in the areas of study is not
reported.
Scientific study has become a piecemeal approach where the duration of
the study is typical of obtaining a doctoral thesis. Continuity is largely
absent. Present-day science consists of collecting disjointed facts that
cannot be repeated and confirmed elsewhere. Experiments do not result in
falsifiable theories. There is no insight.
Studying degradation is not 'sexy'. It does not earn scientific admiration.
We have omitted monitoring the sea for its clarity and sedimentation rate.
A network of simple sedimentation traps on all wharves and some buoys could
have kept us uptodate with the disastrous trends that have been with us
for over fifty years, but which are accelerating steeply now. This has
not been done. It is not even being considered.
Many marine scientists are politically motivated, having traded objectivity
for their beliefs. Science funded by conservation organisations is bound
to give results pleasing them, like that funded by the tobacco industry
pleasing its funders. One does not easily bite the hand that feeds.
The DoC (Department of Conservation, NZ) funded research cannot be critical
of DoC.
Many scientists now earn their living from marine reserves. It has become
a major source of income for many institutions. Only studies related to
global warming exceed this.
A swarm of politically motivated marine 'scientists' is now involved with
marine
reserves (and now also global warming), quoted as a 'growth industry'.
They want marine reserves to work for the environment, science, fisheries,
biodiversity and much more but they fail to see when and why these reserves
don't work. Many work with computers and models to make their point.
These 'scientists' do not take the precaution of distinguishing apples
and pears, and uncritically apply findings about tropical reefs to the
situation in temperate waters. Even there they do not distinguish the special
nature of some spots on the coast. The area around Goat Island for instance,
has always been a special place without equivalent. To compare other places
with it must be done with care. Yet these 'scientists' do not exercise
such care. FAQ.
Nearly all travelling protagonists for marine reserves are not frequent
divers, having no personal appreciation. They are not marine naturalists.
Yet their influence has swayed many and has been decisive for this Government's
flawed policies (NZ is no exception). Consider for instance the long list
of marine research leaders at the University of Auckland: Chapman, Morton,
Miller, Ballantine, Foster, Bellamy, Taylor, Bergquist, Creese, Rees, Costello,
Bowman, Montgomery, none of whom are divers. Is this also the case at other
universities?
So there exist ample reasons to be concerned about the quality of marine
science in the field.
The main issue we contend is the finding that the sea urchin habitat
zone is unnatural and that it disappears in marine reserves which have
had sufficient time to acquire stocks of large predators, particularly
predatory fish (like snapper) and crayfish (rocklobster). We contend that
this is not true and that scientists have taken insufficient precaution
in their experiments. They have also taken insufficient precaution in attaching
far-reaching conclusions to failed experiments. As counter arguments, we
dissect the very research done by these scientists, supplemented by work
done by ourselves and by extensive observations. Since first publication
(2003), a number of overseas scientific publications have supported us.
the ecology of barrens In the study of ecology, the way species interact while depending on
one another, has been a subject of intense speculation. One often encounters
the idea that all species are interconnected by an invisible 'web' where
each is as important as any other ('all things are connected').
However, this idea or model as scientists call it, is entirely wrong.
Since there exists a discernable food chain with producers (plants)
at the bottom, grazers above it and predators above grazers, and many more
grazers than predators, there must be a mechanism that regulates it. The
question is how?
If predators eat too many grazers, they run out of food and die, and conversely
if grazers multiply too much, they will also starve. To explain the 'balance',
there are two models:
top-down control: the predators keep the grazers in check, preventing
them to outgrow their food supply (plants).
trophic cascades: from the top-down control model, it follows
that if people kill the predators, there will be more grazers, and thus
less grass. This is essentially where the snapper-urchin-kelp myth
comes from. It is a bit akin to saying: "when we shoot the bears, the
forest dies."
bottom-up control: the amount of grass determines the number of
grazers which determine the number of predators.
birth control: there are other ways in which populations control
their numbers, and predation (exploitation) is not the only and most decisive
one. In the end, populations adapt to their particular circumstances by
the process of evolution, particularly on islands where habitat size is
limiting.
economics: every population is exploited by others, and the rate
of exploitation depends on how easy it is to find next meal.
extreme events: random extreme events like storms, floods, fires,
droughts, diseases, oil spills have a major and controlling effect on populations.
gradual events: pollution, eutrophication, changing ocean currents,
upwellings, changing climate. Often an unreported gradual event can explain
the effects observed. Eutrophication is the most insidious because it is
wide-spread, cannot be measured and has engulfed most marine research stations
worldwide, while being ignored by many.
We discovered that the first model is wrong. The control
comes from the flow of energy. Thus plants control the number of
grazers who control the number of predators, much the same as in
society where those with the money control those who are paid. The top-down
effect is necessarily much smaller and is best described by the economics
of exploitation (a must-read), where predators are kept in check
by the amount of effort it takes to find food. This also applies to grazers.
The sea in its weirdness adds another dimension to this problem. On
land habitat zones (desert, savannah, forest) are so large that those who
live from it are also living on it. In addition, the offspring
of all terrestrial species are born near their parents. And the
number of offspring is limited to a few. Although this is never mentioned
in books about ecology, these points are of major importance to understand
the sea, where it is often not the case as the habitat forming organisms
spawn profusely (millions of offspring) and their offspring is born somewhere
else. It implies that the genes of survivors are not expressed where the
survivors live. Evolution and adaptation in the sea are thus necessarily
weak. In the sea the narrow habitat zones of the rocky shore are frequented
by predators who do not live there, like snapper. The very narrow
intertidal zone is even visited by grazers who do not live there.
The rocky shore where the urchins live, is but a narrow fringe of the
wide open sea with its vast expanse of sea bottom and water above it. The
sea bream snapper(Sparus auratus) lives mainly over the
sandy and muddy sea bottom, and only very few take residence on the rocky
reefs. It has been shown that most fish specialise in a certain diet during
their lives, and snapper being quite versatile, can be found as predator,
scavenger, crab-and-worm eater and mollusc eater, yet seldom all of the
above. The point I wish to make here is that there are high numbers of
snapper that could arrive from the open sea to wreak havoc on coastal
populations, but very few actually do. And those who could
do harm, are not constrained by a marine reserve. The situation is different
for rocklobsters who live on the rocky shore, but they scavenge more than
predate.
The trophic cascadesmodel (or paradigm as the
authors called it) became popular after a publication (1974) by Estes &
Duggins [1,2,3 and many more] who saw a correlation
between sea otters (who eat urchins) and the giant kelp (which can be eaten
by sea urchins). Thus catching sea otters for the fur trade can have a
decisive influence on the algal assemblages it was thought. This
uncritically became established knowledge and some of the most cited articles
in marine ecology. But although a correlation was seen, a causation
was never established (see below)! So much for trusting marine science,
its practitioners, its peer reviewers and its prestigious journals!
Finally in April 2006 (22 years later!) the trophic cascades paradigm
was tested by Carter, VanBlaricom and Allen [4], showing
that there is no significant increase in algal growth when sea urchins
are removed manually inside the San Juan Marine Reserve, not even over
a longer time period of 2 years. One would
have thought that this experiment would settle the matter, but please note
that their study happened in a sheltered place (San Juan Channel, Washington
State) where storm barren zones cannot occur, while urchin densities were
very low (1 per square metre). This means that the available sea urchins
had no influence on their environment, and removing them of course had
no influence either. The study is thus too weak to disprove the trophic
cascades paradigm.
The
Monterey Bay National Marine Sanctuary has been closely monitored for many
years. The graph shows how sea otter numbers (red curve) increased from
near-zero in 1900 to over 2000 in 2000, proof that they had been hunted
to near extinction. However, recently their numbers have been declining
but why is not known. The kelp harvested here is mainly the giant kelp
Macrocystis
pyrifera (and Nereocystis luetkeana) which prefers sheltered
waters, but even so is erratic in its year to year densities. They are
also easily removed by waves. The blue curve shows total kelp harvest in
all of California, climbing as otter numbers increase, suggesting that
there is a linkage between the two. But there are inconvenient exceptions.
Remember that the influences of market demand and price, regulation, nutrient
supplies and degradation can be decisive. The green curve shows kelp harvested
inside the sanctuary, declining as otter numbers are increasing. Kelp is
harvested by cutting the floating fronds off their stipes. The seaweeds
are used to cultivate abalone, and they are exported to Asian countries.
Monterey Bay was famous for its sardine fisheries wich crashed in 1940-1960,
presumably due to overfishing and pollution.
In the graph also the commercial harvests of red and pink abalone are shown,
both declining steeply around 1970. The green and black abalone are much
less common but also show steep decline (not shown here). Data for urchin
catches are not available. The curves suggest that the Californian coast
and Monterey Bay suffer from severe degradation affecting all species.
tethering urchins to confirm predation Predation on urchins is usually studied separately
by tethering urchins to the substrate. In the laboratory a string is threaded
through the urchin, which causes it to bleed, as well as internal damage.
But the urchin is first kept to recover, before transplanting. The experiment
then shows that urchins inside protected areas disappear faster than outside,
proof that predators remove them. But is this true?
We found that rocklobsters can quite happily walk
over an entire cluster of sea urchins without attacking any. However, when
we damage some by scratching some spines, only the damaged urchins disappear
within days. Rocklobsters are apparently strongly attracted to bleeding
and dying organisms, and nobody can guarantee that a tethered urchin does
not bleed as it attempts to untether itself. By tethering an urchin, it
is not placed where it wants to be and this causes stress. See the
tethering
experiment below. The bottom line is that tethering experiments
confirm that there are more predators inside than outside protected areas,
but not necessarily that these predators (scavengers, really) usually eat
sea urchins. Neither does it prove that a decline in urchins is caused
by predators.
We discovered that urchin barren zones are created by large
storms and that urchins and other grazers just maintain these. Urchin
barrens are thus a normal and healthy habitat, not in any way caused by
humans. We propose to rename them storm barrens, for this is what
they are. Such barrens are defined and restricted by wave propagation laws,
which explain where they are found, where they cannot occur, and how deep
they extend. One of the deepest storm barrens is found on the northern
side of Niue Island where due to extreme water clarity (40-80m viz), the
photic zone extends far beyond diving depth. Poorly occupied by day but
grazed by numerous fish species, the night brings out a true city
of sea urchins in perhaps two dozen different species. Read our account
of Storm barrens in
New Zealand, and underwater images from Niue,
its ecology, index1
and index2 (for large picures, our
CD
is required).
[1] Estes, J A & J F Palmisano (1974): Sea otters:
their role in structuring near shore communities. Science 185: 10858-1060.
[2] Estes JA, N S Smith & J F Palmisano (1978): Sea
otter predation and community organization in the western Aleutian Islands,
Alaska. Ecology 59: 822-833.
[3] Estes, J A & D O Duggins (1995): Sea otters
and kep forest in Alaska: generality and variation in a community ecological
paradigm. Ecol. Monog. 65: 75-100.
[4] Carter, Sarah K & Glenn R VanBlaricom, Brian
L Allen (2006): Testing the generality of the trophic cascade paradigm
for sea otters: a case study with kelp forests in northern Washington,
USA. Hydrobiologia (2007) 579:233-249 (freely available on the Web)
Urchin barren productivity
(the myth of urchin barren unproductivity) Over 200 scientific publications and over 1000 news and opinion articles,
refer to the extremely low productivity of sea urchin barrens cited by
Chapman [1]. Here are some statements:
In reef habitats across the globe - when uncontrolled - sea
urchins have devastated reefs, leaving sea-urchin barrens devoid of
algae. Sea urchin barrens are increasing along the east Tasmanian coasts
reducing
the productivity of the reefs by 80-90%.
Dense aggregations of urchins (or feeding fronts) form along
the margins of kelp beds and destructively graze all upright seaweeds,
creating urchin barrens of much lower productivity, habitat complexity,
and biodiversity.
Sea urchin overgrazing is associated with the transition
of macroalgal-dominated reef habitats to barrens, and such a change results
in a 100-fold decrease in primary production (citing Chapman, 1981
[1]).
Prof. Johnson said the sea urchin had created barren areas
along the East Coast by overgrazing seaweed and invertebrates which had
sustained the abalone and rock lobster fisheries. It really is the
equivalent of taking a bulldozer to a rainforest and clearing everything
to bare earth. Its a major crash in biodiversity and productivity,
with a big impact on important commercial fisheries,
An urchin barren is an area of the subtidal where the
population growth of sea urchins has gone unchecked, causing destructive
grazing of kelp beds or kelp forests. (Wikipedia)
in the early 1970's, grazing by aggregations of sea urchins,
Strongylocentrotus
droebachiensis, completely destroyed subtidal kelp beds in St.
Margaret's Bay, Nova Scotia, leaving barrens devoid of macroalgae.
Destructive grazing of kelps by advancing urchin fronts results
in the formation of barrens, with dramatic losses of productivity and
biodiversity
etc.
There are a number of fallacies in the above statements, beginning with
the productivity of urchin barrens, originating from Chapman [1] who scraped
some alga off a barren and then estimated its productivity: "The
mean standing crop of benthic microalgae was found to be, 2.2 g C m-2
and production estimated as ca 15 g C m-2yr-1 at
8m depth. Most of the primary production of St. Margaret's Bay has been
lost with the disappearing kelp populations." Nobody
of the hundreds who cited this article, exercised care for it was only
an estimate and not actually measured. Then nobody saw that it clearly
had to be nonsense. Imagine your lawn growing only by a cupful of grass
per square metre in a whole year, not to mention that in the coastal sea
there is a large supply of nutrients and moisture!
Admittedly, it is much easier to measure large
seaweeds, by cutting circles from their leaves, measuring width, length
and weighing them and so on, which led to a kelp forest productivity of:
the giant kelp Macrocystis of 0.2 - 2.6 gC/m2/day:
(Robert L. Vadas Sr et al. 2004), or 75-950gC/m2/year
Macrocystis 2kg/m2/year wet weight (), identical
to tropical rain forest?
tropical rain forest quoted generally at 9-11 kg
C/m2/year. (Anderson 1992)
mediterranean mid-littoral up to 10gC/m2/day (Ballesteros
1992)
So there exists a healthy disagreement over the productivity of kelp. But
to claim that barrens are far less productive than kelp forests, overlooks
the fact that pastures (barrens on land) are more productive than forests.
Productivity is not related to the standing stock which usually contains
much woody substance. There are good reasons to expect barrens to be more
productive than forests of brown kelp in edible matter:
the barrens are more shallow and receive more light.
because they are covered in crustose coralline algae with a light reflective
pink colour, plants growing there receive the light twice.
single celled and filamentous algae grow faster than large kelps because
they do not form woody tissue (cellulose cell walls) and can grow as fast
as diatoms do.
green cells are more efficient than brown seaweeds (Phaeophyta).
filamentous algae are more palatable (edible) than brown seaweeds.
ALL the filamentous algae are eaten, whereas LITTLE of the brown macro
algae.
The fact that the haze of filamentous algae covering the rocks cannot be
seen because there are so many grazers who by day and night (24/7) remove
it, does not mean that therefore productivity is also low. The productivity
of barrens can be measured only by first removing all grazers, not just
urchins.
[1] Chapman, A.R.O. 1981. Stability of sea urchin dominated
barren grounds following destructive grazing of kelp in St. Margaret's
Bay, eastern Canada. Mar. Biol 62:307-31 1.
Acrimonious correspondence Our driticism of marine studies done here in NZ and elsewhere has not
been left unnoticed, and we publish it here for you to judge. Please understand
that the criticisms expressed on this chapter are not personal. They are
directed at the research done, the methods and conclusions. More correspondence
in opinion.
A letter from Russ Babcock answered by Floor Anthoni
I was amused, but not altogether happy, to see what you
have to say in your recent posting to the NZMSS list. The style of
writing and the naivety of some of the ecological interpretations you have
made are reminiscent of some of the most outlandish writings of "creation
science" advocates. Do you not care that your musings have made you
an object of ridicule among the scientific community (and increasingly
among the general public)? You should, because they only serve to
discredit the cause of marine conservation and play into the hands of those
who wish to ignore the consequences of continued environmental degradation.
The fact that you continue to do this suggests that notoriety, rather than
real concern for the environment, is the real motivation for your actions.
Such speculation aside, I challenge you to scientifically
refute any of the work you have chosen to highlight in your web pages.
To do this, you will have to take Sam McClatchie's advice, and enroll in
some basic ecology papers at University. You will find that the topics
that concern you have been the subjects of intensive research for some
time. Actually collect some data, display and analyse it, and test some
of your ideas; you might begin to get somewhere. Write up your results
in a coherent form and submit them to review by your peers. If the
work passes scrutiny it might even get published. All of the papers
you have placed (often illegally) on your site have been thru this process,
which is not something that can yet be said about your ideas. Your
claim that "scientific data is missing" smacks more than a little of hypocrisy.
If you devoted half as much energy to constructive enquiry as you do to
your current style of sensational and ill founded exposition I am sure
you would achieve much more positive outcomes for the marine environment
about which you claim to care so deeply.
Yours etc Dr. Russ Babcock
10 Oct 2003
Dear Russ,
I knew you would be unhappy with the marine research
criticised by me on the Seafriends web site, because much of it relates
to the work done by you. I have taken your own work and refuted it on your
own findings, with my observations added, which are there for all to see
and to refute if possible. You must be aware that as long as you are speculating
that the environmental changes you are observing in marine reserves are
related to their long-term 'benefit', you lay yourself open to this kind
of rebuttal. In your letter you mention environmental degradation, but
you don't seem to recognise it in your work.
You (and other scientists) must start by taking each of
the points you disagree with and refute them one by one. I will publish
this on the net to let people make up their minds. In the end, science
does thrive from discussion. I will change what I have written if you are
right and I am wrong. It is as simple as that. Every page on this web site
asks for scientific input and correction. But don't forget that I too have
done my homework and done a lot of studying and research of published scientific
fact. Study the references and 'further reading' to satisfy yourself. Also
don't underestimate my knowledge of New Zealand's seas.
To say that I have made myself the ridicule among the
scientific community, is true particularly with those who profess to be
scientists but are not in their way of thinking. What you fail to recognise
is that I have also gained a lot of respect from those scientists who care
to be challenged. I write about new ideas and pass these on for scientists
to investigate. I have many letters of appreciation from overseas scientist.
Your statement that I don't care for the environment but
am out to gain notoriety, is hurting. Just look at what I have been through
so far in the past 13 years. Can you mention anyone with this amount of
commitment for no pay? It is not easy to be the one noting the emperor
has no clothes, and having the courage to say so. But I carry on because
in the end, we must do the right things for the right reasons at the right
time. We owe it to our children.
So, take a positive step and take some time to comment
or criticise each of the points on the Seafriends web site that you find
is wrong; provide proof; send it to me and I will correct my mistakes.
For if you don't, you have said nothing at all. Invite others to do so
too.
Regards, Floor Anthoni.
Many chiefs
but few indians
An often overlooked problem for small countries with few
people like New Zealand (4 million), is that nearly everyone becomes an
expert in his own field. There are just too few people to support duplicate
experts. In industry and government departments one finds very shallow
management pyramids. In this environment many a novice beginner becomes
manager in as little as two years, the syndrome of a nation of many
chiefs but few indians. People thus quickly progress to a position
of incompetence (Peterson's Principle) or at least escaping criticism from
subordinates, peers and superiors. In a populous state like the USA, this
is much less so. Such syndrome affects particularly the scientific community
where nearly every scientist lives on an isolated island of expertise,
likewise escaping critique from subordinates, peers and superiors.
Specialisation in science has led to small groups or
cliques sharing their experiences, visiting the same conferences while
refereeing one another's papers. In such a close côterie robust criticism
is not delivered for fear of alienating oneself. Whatever the reasons,
it
is deplorable that the scientific literature on marine reserves contains
so much opinion, speculation and flawed research with many citations of
the same.
Own research
Immediately after the mass die-off of the kelp forest inside the Goat
Island marine reserve, we undertook a survey of the extent of the damage.
We were there when dense plankton blooms covered the area between October
and December 1992. We warned the scientists that the kelp would not be
able to survive. Before Christmas 1992 we observed that all the kelp had
already died, and again we warned the scientists of this. In the first
week of 1993 cyclone Oli just finished it off. Not a trace was found of
the entire kelp forest's canopy deeper than 15m. Only the stalks stood
as evidence that a forest was once there. Again the scientists were informed,
but nobody could take a look because of the summer holidays. Only after
these holidays, did scientists discover that the kelp forest had indeed
disappeared.
Scientists were confused finding kelp death both in the shallows and
deeper than 15m but not in between. They could not conceive that the one
had been caused by wave damage whereas the other was caused by lack of
light. As a result they went about their investigations in a less than
thorough way. They did not recognise the importance of such a major experiment
conducted by nature and the possible lessons that could be learnt from
it, and its possible ramifications. This spurred us to do an extensive
survey to establish the precise areas affected and what other lessons could
be learnt. It taught us much about the ecology of the sea coast, particularly
that of kelp and urchins. You can find this report in www.seafriends.org.nz/enviro/habitat/survey93.htm.
Our main findings relating to urchin barrens:
urchin barren zones are found only on exposed coasts
urchin barren zones do not occur in shallow water
barrens do not need to have urchins
barrens are first caused by large storms, then maintained by grazers (urchin,
Cooks turban snail, paua)
Our studies of the marine
ecology in Niue confirms this hypothesis as hurricanes create very
deep barrens only on one side of the island (2004, 2005).
there are more grazers of young kelp than thought, including planktivorous
fish.
Also confirmed in Niue.
the disappearance of the urchin barrens at Leigh is caused by the mass
dieoff of the kelp forest.
It is important to distinguish barren patches caused by urchin grazing
from the contiguous barren habitat zones. Even where no barren zones are
expected, one may find an occasional barren patch.
An important result of our survey was the measured habitat zoning/exposure
map which is representative of north facing exposed to sheltered rocky
shore on the east coast of the North Island.
Our version of the urchin
barrens story
In the period that the kelp forest disappeared, we were fortunate that
we were able to make many dives around Leigh and further out. Because of
our school programmes during which we guide students snorkelling the Goat
Island Channel, we were frequently in the sea from September to May every
year. Dense plankton blooms are not welcome because it hampers the programme
and reduces the 'wow' experience of the school visit.
Because
the plankton blooms between October and December 1992 were exceptionally
dense, a few exploratory dives were made to observe plant life in the dark.
At the peak of plankton blooms, visibility was less than 2m at the surface
and the light at 20m depth was so poor that the sandy bottom could hardly
be distinguished from the dark rock. It was darker than a moon-lit night.
In November the kelp turned dark brown and started to look like wet kraft
paper on woody stalks. By mid December, their crowns fell apart when touched
but in shallower water the kelp appeared entirely unaffected. It was clear
that a major event had happened, and we informed the Marine Laboratory
accordingly. The photo shows what the dead kelp forest looked like.
In
the first week of January 1993 tropical cyclone Oli struck, and when diving
became possible again, we saw that the whole kelp forest below 15m had
disappeared. In the months following, the kelp died back further to effectively
disappear where once the forest stood. Some shallow water dieback was also
evident but this was caused by the waves of cyclone Oli. The second photo
shows what an extensive sea urchin barren looked like (June 1995, Waterfall
Reef), with a cluster of sea urchins in the foreground and a leatherjacket
(Parika
scaber) above. Note that the kelp had not yet invaded the barrens.
In March 1993 we did an exploratory study in order to prepare a more
extensive one, which we did in the winter of 1993 (Aug-Sep). The results
of this study you can find on this web site in /enviro/habitat/survey93.htm.
The above two maps are important to understand why the urchin barrens
theory is a myth. The first shows where the 91/92 kelp deaths occurred,
but more importantly, the second shows where the 92/93 kelpbed death occurred
and to what extent. The areas marked 4 and 5 lost 80-100% kelp cover, and
all these areas recovered the way of Goat Island, with loss of urchin habitats.
Yet these are not marine reserves. Here is what happened to their kelp
and urchin habitats:
This
diagram depicts our observations in the years preceding and following
the kelpbed death events. On the left the situation where the kelp bed
was removed twice, first partially in 1992, then fully in 1993 (Little
Barrier, Simpsons Rock, Arid Island). On right the situation where the
kelp was removed once, but completely.
In 1991 both areas had a mixed age kelp forest. In 1992
part of the kelp forest disappeared in some areas, followed by rapid recruitment
and full regrowth because grazers had plenty of food in the remaining stands
of kelp. The urchins stayed in their barrens. By late 1992 the kelp forest
had almost complete coverage.
In Dec 1992/ Jan 1993 the kelp died, again because of
dense plankton blooms. Recruitment was immediate and intensive in the areas
where the kelp once stood but not outside these. However, all regrowth
was eaten by the remaining grazers, which included fish. The urchins strayed
from their barrens into the deep, leaving their barrens insufficiently
grazed.
Successive waves of recruitment but much less intensive
followed. Kelp settled slowly into the urchin barrens and the areas where
the kelp once stood. This time it was successful and it took over the urchin
barrens. The full takeover took several years until the kelp reached maximal
growth rates. Isolated reefs like Floors Reef and Leigh Reef took until
1998 before they were recovered fully. These have no urchin barrens, yet
grazing there was very intensive.
We also noticed massive urchin deaths and poor recruitment,
which affected the process.
From the scientific research dissected on this page, it can be seen
that scientists were not aware of the above, even though they were in possession
of our Hauraki Gulf Marine Survey 1993 report.
The changes in community structure
by top-down control (the urchin barrens theory) was a reverberation
of the 1993 kelpbed death. It was caused not by the beneficial effect of
no-take protection but by degradation! The urchin barrens theory is a myth.
Recurring mistakes
Some of scientists' mistakes keep returning in many of their studies. Rather
than elaborating on them each time as part of their research, we discuss
them here.
The Goat Island marine reserve, located under the Marine Laboratory of
the University of Auckland was created specifically for doing marine research.
Not surprisingly, much work has been done here, making it one of the best
studied areas in this country. Being also the first marine reserve, much
of these findings are explained by them found inside the marine reserve
rather than outside. Now that protagonsists for marine reserves need more
reasons for having more of them, scientists are pressured to do research
to prove that marine reserves 'work'. In doing so, they often compare inside
with outside, taking the Goat Island marine reserve as the inside dataset.
However, this is an oft repeated mistake, ignoring how special the environment
around Goat Island really is. Unfortunately for this kind of research,
Goat
Island is not at all representative of the surrounding coast. Both
fish and visitors know that, but scientists keep ignoring it. Scientists
should take more care when using Goat Island for comparative studies.
It
repeatedly results in overestimating the number of fish and crayfish in
a typical marine reserve.
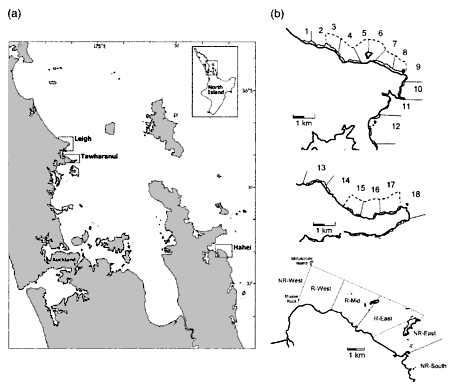
The map above is a composite of three maps drawn by Land Information
(LINZ), which was the Department of Lands and Survey. The data for this
map was obtained by a team of divers under leadership of Dr Tony Ayling
in 1977. Click on the map for a page-sized one. The depth of the rocky
shore ranges from 15m in the west to 23m in the east with a sandy bay about
one quarter from west ending in a shelly beach.
The habitat zoning can clearly be seen. The shallow dark brown zone
consists of tough bladder weeds (Carpophyllum spp). The grey/blue
zone corresponds to the urchin barren zone. Notice the large area of submerged
rocky shore around Goat Island, which is unique. Because of its gentle
slope, most of the barren zone is also found here. Also unique are Goat
Island's areas of shelter preferred by fish, its extensive crayfish habitat
and shellfish beds, caves and broken rock (large boulders). To the west
of Goat Island the substrate consists of layered mudstone; to the east
of (broken) hard greywacke. Note how the shores to the west and east become
more representative of the coast in this region, having a narrow rocky
substrate. Further west extends the 25km long Pakiri Beach, which makes
Goat Island the first rocky outcrop with shelter and currents, like
an oasis at the end of a long stretch of desert. There is no place
like Goat Island in a vast region around.
Scientists also do not take sufficient care in comparing apples with
apples on the choice of their transects and areas of study. They believe
too much in the impartiality of dice. Yet transects and areas of study
must be chosen carefully to match exposure, aspect (orientation), depth
and isolation (sand, crevices, walls). Otherwise too much spurious data
will spoil the experiment.
Scientists view snapper (Pagrus auratus) and crayfish (Jasus
edwardsii) too much as predators rather than scavengers. These
animals are strongly attracted by stressed organisms such as damaged sea
urchins
(Evechinus chloroticus), shellfish and other. Likewise the
common urchin (Evechinus chloroticus) is strongly attracted to stressed
kelp.
Scientists make no mention of major oceanographic events happening
during their experiments (or just before), events that could have influenced
the outcomes of these experiments. Chronology, mentioning the precise start
and end of the experimental period is therefore important. In their papers
they fail to report the time of study relative to large events like cyclones,
kelpbed deaths, large urchin mortalities and the disappearance of crayfish.
Scientists are so keen to show overwhelming benefits arising from
marine reserves that they lose objectivity. It has led to nonsensical
conclusions as the reader may see for himself. The fact that much research
was funded by the Department of Conservation who manages reserves and whose
task it is to create more, did not help - one does not bite the hand that
feeds. As a result, politics have entered science. Read also our extensive
myths
and fallacies section.
The baited underwater video camera (BUV)
is a highly questionable measuring device since it aims to influence the
quantity measured rather than minimising its influence as is required for
any other scientific measuring device. Not only does it bait the camera,
but it also offers an additional pilchard as food! Such a method grossly
overestimates the quantity measured and should never have been attempted
as a scientifically acceptable measuring device. But it can be used qualitatively
to say that at some place there are more or less fish than at some other
place. Scientists have never proved its linearity (as in a voltmeter) for
quantitative measurements. [in a linear instrument, there is a one-to-one
correspondence between the measured and actual quantities]. For a description
of the BUV method, see further on this page and in our
Frequently
Asked Questions about marine reserves.
Conclusions By exposing a number of scientific experiments
on this page, we have shown that marine ecological research in New Zealand
has not matured. Marine scientists have drawn major conclusions from failed
experiments while not taking sufficient care in comparing apples with apples.
Rather than following the traumatic events of 1992/93 with an interest
in learning from them, scientists have been only too keen to explain ecological
changes in terms of the success of marine reserves rather than due to their
failure (degradation, kelp death, mass mortalities, poisonous slime).
It did not help that scientists still do not understand the ecology of
the rocky shore in their areas. Neither did it help that the research was
funded by an ideologically motivated political bureaucracy whose better
judgment is clouded by a pervasive zeal for creating more marine reserves.
Our own research adds a refreshing turn to the issue, filling in the
missing bits while providing new insight into the rocky shore ecology.
Although it is not our task to do marine research, it is nonetheless the
task of the public to expose faults and fallacies where scientific self-policing
fails. We are waiting for the scientific community in Leigh to correct
its mistakes, by looking at the bloom-affected areas around the outer Hauraki
Gulf, in order to arrive at the inescapable conlusions:
the snapper-urchin-kelp hypothesis is a
myth. marine reserves do not cause sudden ecological
changes after a long period of stability. urchin barrens are created mainly by storms. to prove that coastal marine reserves are
working is wasting time and money. marine research should focus on studying
the causes and effects of land-based pollution and degradation.
The extent of die-back of
the kelp Ecklonia radiata in the Cape Rodney to Okakari Pt Marine Reserve Advice to the Department of Conservation, June 1993
R C Babcock and R G Cole University of Auckland Leigh Marine Laboratory
[our comments in blue]
Summary High levels of mortality have occurred in beds of the kelp Ecklonia
radiata in a region stretching from Whangarei Heads south to the Okakari
Point to Cape Rodney Marine Reserve and east to Great Barrier Island. The
area most heavily affected appears to be centred on the marine reserve
where mature kelp plants are virtually absent from many areas, and densities
overall are an order of magnitude lower than in 1991.
In
our own study we excluded the areas north of Whangarei after a preliminary
survey in March 1993. It allowed us to concentrate more on the areas surrounding
the outer Hauraki Gulf. Because of our extensive spatial data, we can say
that the bloom and its mortality did not centre on the Goat Island marine
reserve. See our map on right. The area around Port Fitzroy on Gt Barrier
I is rather patchy, possibly because this harbour has its own plankton
ecosystem and may have been spared some of the bloom's ravages. Also unaffected
water flows north and out of the Hauraki Gulf past these areas.
Mortality is patchier in other areas such as Great Barrier Island. Recovery
of affected populations is already underway with high levels of recruitment
having taken place at the marine reserve between January and May 1993.
Full re-establishment of kelp populations should proceed over the next
year, unless recruiting kelp become susceptible to mortality in the post-juvenile
phase. The causes of the mortality are not yet known but all hypotheses
put forward to date have related to natural environmental processes and
human impacts are not implicated.
It is always surprising how the human
factor is reasoned out of the equation so quickly. If something is unique
or cyclical, then humans are not at fault, it is reasoned. But what caused
the concentrations of nutrients necessary for this plankton bloom build-up?
When is a unique event just the first of many to come? When is a deteriorating
trend from moderate kelp die-off in 1982 to a large one in 1992, recognised?
When are such events admitted to be degradation of the environment?
Fig 1. Kelp density before and after mortality events. Mean
density of Ecklonia radiata in the Cape Rodney to Okakari Point
Marine Reserve based on five 1m2 quadrats at each depth. Depths are maximum
depths for each depth strata (e.g. 3-6m, 6-9m, 9-12m)
These two data points are accidentally
the only ones showing the situation before and after. The top two curves
of the before situation, show that the kelp forest is found deeper than
9-12m. After the plankton blooms the deep kelp disappeared, leaving the
shallow kelp unaffected. Note how kelp diminishes with depth in the bottom
diagrams.
Local mortality patterns The proportion of kelp populations damaged or intact varied considerably
among sites within the marine reserve (Fig 4), though all sites had low
densities of intact plants. The proportion of intact plants was relatively
high (over 50%) at Marimo and Point, but was much lower elsewhere. For
example Waterfall and 'D' the mortality process appeared to be still active.
The proportion of the populations contributed by recruits was also variable
with North Reef and Lookout having the highest levels. Despite the overall
variability, highest levels of damage were consistently seen in the deepest
levels at each site.
Fig 4. Local variation in relative kelp mortality, May
1993. Data presented are proportions in each damage category based on video
transects and they represent all depth categories.
The authors made a major error of judgement
by including recruitment, which happened after the kelpbed death,
in their results, rather than dealing with this separately. The pie diagrams
should also have included barren areas without kelp. In this map a shallow
shore like Marimo seems to have sustained little kelpbed damage whereas
in reality it does not go deep enough to have a substantial kelp bed. The
map cannot be interpreted without much local knowledge. Had the authors
displayed the amount of kelp canopy left as part of the natural kelpbed
zone, it would have been very clear that the kelpbed death is very consistent
along the shore. The method also becomes useless in areas or zones which
have been hit by a kelpbed death in two successive years, see below.
Regional
mortality patterns A survey of sites between Cape Karikari in the north and Kawau Island
in the south, and offshore to western Great Barrier Island, revealed that
in this region kelp mortality was restricted to the outer Hauraki Gulf
area. Sites at Whangarei Heads, Okakari Pt to Cape Rodney Marine Reserve,
Little Barrier Island and Great Barrier Island were all affected by recent
large-scale mortality of Ecklonia. However, other sites at Gt Barrier
I were unaffected, as were sites at Takatu Peninsula and Okakari Pt.
This observation agrees with our findings.
What confused the authors is that their method could not distinguish between
sites that had been devastated twice, such as some of the Gt Barrier sites.
The authors have not taken due caution excluding sites with shallow sand
bottoms such as at Okakari Point and some at Takatu. The authors did not
make a distinction between shallow and deep kelp death.
Thus within the region where mortality is evident there was variation in
the level of mortality between sites as well as at local scale within sites.
This local scale variability was evident over short distances with the
first two sites for Gt Barrier Island, being separated by only 100m. See
notes before.
The pattern of highest mortality in deep water was not always seen
outside the marine reserve. The affected site at Gt Barrier I was suffering
high levels of mortality, with over 50% of plants present suffering reduction
or total loss of laminae (excluding recruits), yet the most seriously affected
areas at this site were in shallow water.
In our surveys we identified the seemingly
contradictory site at Gt Barrier I as a marginal case where the kelp jaundiced
but recovered. It was then destroyed in shallow water by the force of Cyclone
Oli. The author's method is not suitable for sites with complete loss of
kelp and no recruitment, yet many such places could be found. Although
the Gt Barrier I site seems inconsistent, it is consistent with nearby
sites. See also notes below.
Recruitment Very few recruits of Ecklonia were detected in surveys of the
marine reserve during January. During March numerous recruits were noticed
in parts of the reserve and by May high levels of recruitment were evident
at most sites. . .
The recruits should have been dealt with
separately since they did not exist before the plankton blooms.
Possible causes of mortality Several scenarios have been proposed in attempts to explain the mass
mortality of Ecklonia radiata in the region under consideration. There
are three basic hypotheses:
Pathogens . . The patchy nature of the mortality at other locations such as Gt
Barrier I is consistent with this hypothesis, rather than a broad-scale
physical phenomenon. . . [viral tests] . . The possibility of viral involvement
is still being actively pursued but the presence of agar in algal tissues
has hampered progress since it interferes with techniques used as assays
for viral activity. . .
Every marine organism with reduced health
runs the risk of attack by decomposing organisms like viruses and bacteria.
To find these pathogens on dead and dying kelp is akin to finding fungi
on rotting apples. What is pathetic is that the authors have been warned
before December 1992 that the kelp forest stood dying and their fronds
drooping like wet kraft paper. It is notable that after Cyclone Oli not
a trace was found of the masses of dead canopies. They seemed to have vaporised.
They did not have any food value left in them. Pathogens cannot achieve
a kelpbed disappearance within several hours or even days.
Environmental factors . . . The first general hypothesis is that mortality is simply and
directly related to low temperatures, an idea consistent with the gradual
onset of the mortality during 91-93, but at odds with the observation that
plants in shallow water were unaffected, and also with the lack of mortality
at points further north.
Ecklonia is found all around the
North and South Islands, tolerating temperatures between 6 and 23 degrees,
and mortality was abrupt, rather than having a gradual onset.
Phytoplankton blooms have also been suggested as a possible causative agent
since they would reduce light available to the kelp for photosynthesis.
The mortality of deep water kelp first in the reserve is consistent with
this hypothesis, as is the regional distribution of the mortality, since
the plankton bloom of late 1992 was most intense in the Hauraki Gulf between
Whangarei Heads and Great Barrier Island. The mortality of shallow-water
kelp in some locations, as well as the continued progress of mortality
long after phytoplankton blooms had dispersed, contradict this hypothesis,
unless the energy debt is so great that plants cannot catch up despite
improved conditions.
The deep kelp death was without any doubt
caused by lack of light. Not only have we seen it happen, we have
also predicted that it would happen! What the authors don't realise
is that the shallow kelp death was caused by wave action. The authors are
rather naive about the nature of life and survival after being exposed
to a gradient of a lethal agent. The Hiroshima and Nagasaki atom bombs
caused immediate deaths but also some forty years later. Even on the way
to recovery, an organism remains susceptible to other attacks.
Demographic cycles Mortality of stands of Ecklonia radiata have been reported in
the past, originally in 1973-74 by Don (1975) around Little Barrier Island,
and subsequently by C Battershill and A McDiarmid at the marine reserve
in 1983-84 (pers. comm.). The estimated longevity of Ecklonia radiata sporophytes
is around 8 years, about the same order of time as the interval between
previous mortalities. Since the presence of an intact kelp canopy inhibits
the establishment of recruits in the understory it may be possible for
a cohort of plants to dominate the benthos until they become senescent.
. . . The demographic hypothesis is not inconsistent with lower mortality
in shallow waters since the age structure of populations in these frequently
disturbed habitats will be more even. . .
Although the above reasoning is valid,
the authors and many before them have never looked at the age composition
of a natural kelp forest. There is talk about the number of plants per
square metre or the amount of cover but the age composition is nevere mentioned.
Thus the transition from a multiple-aged forest in 1991 to a single aged
forest in 1994 has been left unnoticed.
Conclusions Kelp mortality is limited in extent and is seemingly most severe in
the outer Hauraki Gulf region. Within the affected area mortality is patchy,
but is most common below 10m. Populations have been recruiting rapidly
and should recover over the next one to two years unless the cause of mortality
(which has not yet been identified) persists.
The effects of the kelp mortality on other organisms on the reef are
not readily predictable, but appear to be of considerable potential importance
to energy flows, growth rates and behaviour of grazers, and the population
characteristics of fishes. Insufficient long-term information exists to
judge the magnitude of the effects which may derive from this event, or
the significance of the mortality to the reef environment.
The authors give themselves advanced
warning that after a major event, major changes could follow. Yet they
have not taken heed as some of the following papers show how they interpreted
the disappearance of urchin barrens as a beneficial effect of aged marine
reserves (due to the presence of large predators), rather than being related
to the 1993 kelpbed death (degradation).
Changes in community structure
in temperate marine reserves Russell C Babcock, Shane Kelly, Nick T Shears, Jarrod W Walker, Trevor
J Willis
Mar Ecol Prog Ser 189:125-134 (1999)
Leigh Marine Laboratory, University of Auckland
P O BOX 349, Warkworth, New Zealand.
[blue text is ours]
The authors studied two 'old' marine reserves
in northeastern New Zealand, at Goat Island (established in 1975) and Tawharanui
(established in 1982), in order to detect whether changes in protected
predator populations had resulted in other indirect changes to grazers
and consequently to algal abundance. They conclude that this is indeed
the case and that they found more urchin barrens in the protected areas.
They also did urchin tethering experiments which
prove that predation in marine reserves is higher than outside. The authors
asked themselves the following questions, which they answered affirmatively:
has the abundance of predators in protected areas increased?
YES We agree.
has the abundance of invertebrate grazers decreased?
YES We agree.
are changes in grazer abundance caused by predator abundance?
YES We do NOT agree.
has the abundance of macroalgae increased? YES But
not for the stated reasons and not gradually.
are changes in the abundance of macroalgae caused by changes in grazer
abundance? YES To
a minor degree.
Our main objections to this research are
It conveniently ignores other marine reserves
outside the kelpbed death region, taking the only two marine reserves where
barren zones disappeared.
It ignores a previous study by the same author(s)
which gave evidence that there is no difference between inside and outside
in marine reserves.
It does not prove the disappearance of whole urchin
barren zones.
The method used is that of a sledgehammer, by
taking measurements on incomparable coasts, then shaking the results in
a statistical mixer and interpreting these results as proof.
Since the original data have been buried in the
statistical mixer, they cannot be verified.
We cannot verify how much the Goat Island dataset
dominates all results.
The research has been set up to prove a political
objective, not to learn more of the undersea ecology.
This research was conducted after one before it,
comparing five marine reserves, failed. This one worked because both reserves
suffered substantial kelpbed death in 1993.
(having mentioned that snapper and crayfish are both
more abundant inside the reserves) . . Consequntly, kelp forests
were more extensive in 1998 than they were at the time of reserve creation
(1977). Urchin dominated barrens occupied only 14% of available reef substratum
in reserves as opposed to 40% in unprotected areas. These changes in community
structure, which have persisted since at least 1994(false),
demonstrate not only higher trophic somplexity than anticipated in Australasian
ecosystems but also increased primary and secondary productivity in marine
reserves as a consequence of protection. Trends inside reserves indicate
large-scale reduction of benthic primary production as an indirect result
of fishing activity in unprotected areas.false
The
1977 date is misleading because the situation stayed stable until the
large kelp dieback. Not a word about the large-scale kelp forest die-off
in 1993. The 1994 date is wrong since kelp recruitment failed that
year due to hungry grazers of all kind.
The conclusion is not at all
substantiated by the research done (see below). Large-scale disappearance
of urchin barrens was caused by the 1993 disaster. This aerial photo taken
in April 1999, shows the main barren area of the Goat Island marine reserve.
Notice how the kelp is beginning to invade, but most of the invasion happened
suddenly a year later. Scientists maintain that it was a gradual process
that began before 1980, which is proved false by this photo.
This
location map shows in dashed lines the countours of the Goat Island marine
reserve in the north and the Tawharanui marine park in the middle. The
numbered areas are the places where measurements were taken. The shaded
circles are the places where lobster abundance was measured.
Methods The study areas were the two oldest marine reserves in New Zealand,
the Cape Rodney to Okakari Point Marine Reserve (hereafter the Leigh marine
reserve, 518ha, established in 1975) and Tawharanui Marine Park 15 km to
the south (350ha, established 1982). Non-reserve reference sites were located
on similar areas of coast adjacent to the reserves.
It is misleading to call these the two oldest
marine reserves, even though they are. Why are the Poor Knights (1981)
not included? Why is this not explained? Our objection is that north facing
coasts (exposed) cannot be compared with south-east facing coasts (sheltered).
The outcome of the results of this study could be interpreted as 'north
facing slopes have lost their barrens' rather than 'marine reserves did'.
Another problem of the choice of study site is
the choice of areas 1, 2, 3, 13, 14 and 15. These shores are too shallow
to have urchin habitat zones. They are bound to cause unnecessary noise
in the data.
Sampling effort was spread evenly across both reserve and non-reserve
areas by dividing them into a number of sites and then sampling haphazardly
within them. This was done to avoid biases that might be associated with
human activities in the Leigh marine reserve, (e.g. fish feeding, c.f.
Cole et al 1990) . . . .
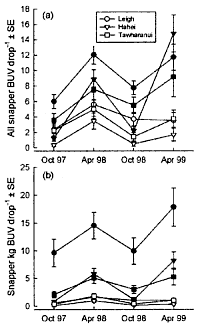
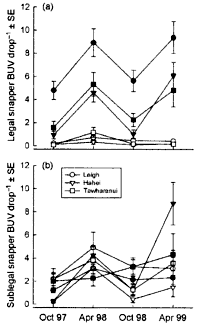
Fish abundance Estimates of predatory fish abundance and size were made during October
and November 1997 using remotely-deployed baited video stations. A vertically
oriented video camera was mounted over an enclosed bait consisting of four
whole pilchards (Sardinops neopilchardus) with one additional pilchard
tied externally to the food container. . . . .
The baited video camera method exaggerates.
See comments about the BUV in recurring
mistakes.
Spiny lobster abundance Lobster surveys were conducted at two sites within and two sites outside
each of the reserves. Sites at Leigh and Little Barrier Island were sampled
in April, and Tawharanui and Kawau Islands in October of 1995. At
each site five haphazardly placed 50x10m transects were surveyed giving
a total of 80 transects covering 4 ha of sea floor. . . . . All sites were
dominated by laminarian and fucalean kelp forests and urchin zones (Evechinus)
at shallower depths.
The timing of these surveys is at odds
with the time of the experiment (1998), since the lobsters were counted
in 1995 when all sites were still having urchin zones. During the time
of the main experiment, this was no longer so. Furthermore, between June
and October 1998, 85% of all crayfish walked out of the marine reserve
due to a prolonged period of heavy rains and sediments (personal observation).
Not a word about this in this paper. Note also that the Little Barrier
Island survey was not part of the 1998 study area, although the areas surveyed
are more similar to the marine reserves they are compared with. The graphs
below must therefore be interpreted with care, also because the results
of each reserve are not shown separately. Leigh may for instance swamp
the data from Tawharanui. Why was it done this way? Our suspicion is that
Tawharanui has a very low crayfish density.
The diagrams show the densities of snapper
(left), as counted with the baited camera and that of crayfish (right)
counted visually.
Algal abundance To test whether urchin-dominated rock-flats (urchin
barrens) were less extensive inside protected areas, we conducted
a series of measurements on 65 transects at 21 sites in and around the
two reserves in December 1997. The sites used were the same as those for
the estimates of fish abundance and an additional three transects were
laid at Kawau Island, 2 km to the south of Tawharanui. . . .
As mentioned before, the SE facing coast
cannot be compared with the north facing reserves, since it is more sheltered
and more shallow. The sites near large beaches are similarly incomparable.
More caution should have been exercised in choosing the survey sites.
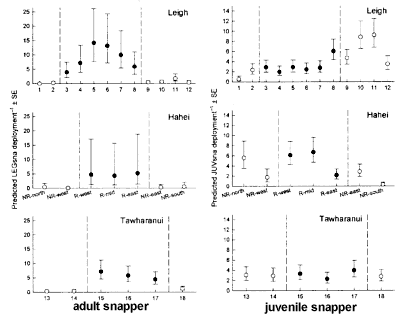
Temporal changes in the density of kelp
and urchins in the Leigh Marine Reserve. Measurements of the abundance and size of kelp
and urchins made in prescribed areas of the Leigh reserve in 1977-78 ("permanent
quadrats" Ayling 1978) provided a direct basis for assessing temporal change.
These data were obtained in a range of habitat types using haphazard 1m2
quadrats within 26 defined 100m2 areas. The same measurements were repeated
as close as possible to nine of these areas in 1994 and fourteen of them
in 1996, using the detailed maps provided by Ayling (1978). All of these
areas were classified by Ayling (1978) as either kelp forest or rock-flats.
Samples from 1994 and 1996 were estimated to be within 20m radius of the
original 1978 areas, and were in the same depth range as Ayling's areas,
as indicated on his habitat maps (Ayling 1978) . . . .
It is a blessing to have permanent quadrats
for monitoring how the environment changes with time. However, Ayling's
maps were used and these have proved to be less accurate. It would have
been more helpful to establish new 'permanent quadrat' sites with accurate
GPS co-ordinates. The dates of these surveys are again not concurrent with
the 1998 date of the main survey, and particularly 1994 is questonable
in relation to the 1993 kelp death. However, the data reflects what can
be seen with the untrained eye.
Primary productivity Estimates of primary productivity were calculated
based on habitat maps (Ayling 1978) and comparisons between our 1996 data
and the data provided by Ayling for the density and population structure
of Ecklonia in kelp forest, rock flats and shallow mixed algal habitats.
. . . For each of these habitat types the primary productivity of Ecklonia
sporophytes was estimated according to Novacek (1980) who obtained size-specific
productivity measures for individual plants. . . [the plant biomass was
measured by measuring stipe diameter] . . Using published values for primary
productivity of temperate crustose coralline algae (CCA) (Littler &
Arnold 1982), the productivity of CCA dominated rock flats was also calculated
on a per-unit basis. . . .Data for each prescribed area in 1978 and 1996
were averaged for each habitat type and used to calculate primary productivity
of the main reserve habitats. . . we estimated that the total area of rock
flats in the reserve has shrunk by an order of magnitude from 31.4% to
3.2% of the total available reef area. . . Overall, the total primary productivity
of the rocky reef habitats we examined increased by 58% from 278 ton to
439 t dry weight per year.
(The analysis is waiting for a copy
of Littler & Arnold's and Novacek's publications).
It would have been more helpful to have measured the productivity in-situ,
since using other people's data from other places in the world, and from
so long ago, is highly questionable. It is not clear precisely what productivity
has been measured. Is it the production of algal biomass? That certainly
does not follow ecological concepts of productivity. It is often better
to estimate the biomass of the grazers because kelp makes much indigestible
biomass. Kelp has very few grazers, which includes fish. Its fronds are
often found rotting back for lack of grazers. Broken fronds are caught
and eaten by sea urchins.
One cannot compare the 1997 kelp forest, consisting
essentially of one year class, with that of 1978 which consisted of many
year classes (personal obs).
The most disturbing aspect of this part of
the research is that it is at odds with ecological principles. The
productivity of micro algae growing on the crustose coralline is much higher
than that of macro algae because of their much smaller size and much higher
growth rate. Furthermore, urchin barrens experience more light and the
light colour of the CCA reflect light back, thus enhancing productivity
further. Even measuring the biomass of grazers is fraught with problems
since urchins spend most of their energy producing spawn.
Reader please note that CCA (Pink paint) is like
living stone, hard and inedible while growing only very slowly. No surprise
then that its 'productivity' is very low. However, the barrens' productivity
comes from fast growing green hairy algae growing on top of the ink paint.
Yet this was not measured. This is akin to claiming that the productivity
of grassland equals that of bare land.
In the discussion we find many nonsense statements: .. . In contrast, all areas that were classified
as kelp forest in 1978 remained stable as kelp forest in 1996. The
author omits to mention that they disappeared completely in January 1993.
This
was despite substantial episodic (sporadic,
at erratic intervals, but there was only one event)
mortalities of Ecklonia during that time (Cole & Babcock 1996, Easton
et al 1997). We propose that higher predation upon and mortality of the
urchin Evechinus chloroticus inside the Leigh reserve is the cause
of the observed changes in community structures within the Leigh and Tawharanui
marine reserves.
Note that other research has been done
on the predation of 'tethered' urchins, see further on this page. The authors
do not mention mass urchin mortality in the 1994-1997 period even though
they studied this as well. But the most worrisome aspect of this research
is the absence of a time study of the populations of the major players,
snapper, crayfish, kelp and urchins. Had they done so, they would have
seen that the 1993 kelp die-off event was the cause of all instability
observed later, including urchin deaths.
Our results also suggest how no-take marine
reserves can change benchmarks for environmental and fisheries management.
This statement is for the benefit of DoC.
The recovery of kelp populations subsequent to
protection indicates that fishing activities on New Zealand's northeastern
coast have had ecological impacts far beyond the target species. These
effects, though indirect, are just as striking as those caused by fishing
practices such as trawling, that remove or destroy conspicuous components
of fished habitats (Rothschild et al. 1994, Dayton et al. 1995, Watling
& Norse 1998). Perhaps more importantly in the case of kelp forests,
these conspicuous components are also major primary producers (Mann 1973),
whose contribution to detrital food webs is central to ecosystem function
and diversity (Duggins 1980). . . . (sigh)
. . . The corollary of this (higher
productivity) is that benthic primary productivity
in areas outside of reserves is much lower than it was prior to intensive
fishing. . . . Kelp forests in northeastern New Zealand are known to have
far higher rates of secondary production than do rock-flats habitats (Taylor
1998), supporting the suggestion that these effects are likely to be broadly
felt throughout the ecosystem. (awaiting
arrival of Taylor's publication)
These observations indicate that the efficient
management of coastal fisheries can no longer ignore the consequences of
fishing on the wider ecosystem and attempts are now being made to understand
the benthic ecosystems in terms of multi-species fisheries models (McClanahan
& Sala 1997). No-take marine reserves represent a large scale ecological
tool that can play an important part in the development of such models.
Unfished areas provide a way of assessing ecosystem function and avoiding
the 'sliding baseline' phenomenon, in which expectations of what is natural
are reduced because may of the original components of the system are reduced
or absent (Dayton et al. 1998). . .
Far-reaching conclusions for what
is in essence an experiment that measured the sliding baseline effect of
degradation, triggered by the kelpbed death in 1993, and did not provide
significant results! (Sigh again)
The myth spreads far and
wide quickly, often mis-quoted!
Palumbi S R 2002, Marine reserves: a tool for ecosystem
management & conservation, Pew Oceans Commission, USA.
Although the author identifies
surprise: p30: "Research within reserves continues to generate surprises
about how marine ecosystems function (Babcock et al 1999)",
he asserts on p24:
"One of the most dramatic reserve effects occurs when
habitat protecion leads to a wholesale ecosystem shift. On the coast of
New Zealand, where fishing for spiny lobster (Jasus edwardsii) has
been severe, urchin populations have exploded and kelp almost disappeared.
Halting the exploitation of lobsters in New Zealand marine reserves brought
the kelp back, along with the fish that inhabit kelp forests, reconstituting
a whole ecosystem (Babcock et al. 1999). In this case, lobster exploitation
cost the marine ecosystem much more than this one species, and protection
restored the ecosystem by limiting biological habitat destruction caused
by overabundant urchins. This is a simple case because only one pivotal
species, the spiny lobster, is present, and it could be argued that single-species
management would do the job as wel. . . " [sigh] National Research Council, 18 authors. Marine Protected
Areas: tools for sustaining ocean ecosystems. p129:
In New Zealand such trophic interactions
impacted even more of the food chain. Because fish are the major predators
regulating the population of sea urchins, closing areas to fishing converted
them from urchin-dominated algal 'barrens' to kelp forest (Babcock et al,
1999) . . . . Studies in the Leigh Marine Reserve in New Zealand indicated
that target fish species can act as keystone predators whose removal causes
strong shifts from sea urchin barrens to kelp-dominated ecosystems (Babcock
et al 1999).
Marine reserves demonstrate
top-down control of community structure on temperate reefs Nick T Shears. Russell C Babcock
Springer Verlag May 2002
Replicated ecological studies in marine reserves and associated unprotected
areas are valuable in examining top-down impacts on communities and the
ecosystem-level effects of fishing. We carried out experimental studies
in two temperate marine reserves to examine these top-down influences on
shallow subtidal reef communities in northeastern New Zealand. Both reserves
examined are known to support high densities of predators and tethering
experiments showed that the chance of predation on the dominant sea urchin,
Evechinus
chloroticus, within both reserves was approximately 7 times higher
relative to outside. Predation was most intense on the smallest size class
(30-40mm) of thethered urchins, the size at which urchins cease to exhibit
cryptic behaviour. A high proportion of predation on large urchins could
be attributed to the spiny lobster Jasus edwardsii.
Predation on the smaller classes was probably by both lobsters and predatory
fish, predominantly the sparid Pagrus auratus. The density of adult
Evechinus
actively grazing the substratum in the urchin barrens habitat was found
to be significantly lower at marine reserve sites (2.2+-0.3/m2) relative
to non-reserve sites (5.5+-0.4/m2) (a factor two).
There was no difference in the density of cryptic juveniles between reserve
and non-reserve sites. . . .
Top-down regulation of biological communities has been a focal point
in ecological theory (Hairston et al 1960). This is ironic, given the efficiency
with which humans have themselves harvested the large-bodied animals which
may be responsible for the top-down control of ecosystems, in many cases
to extinction (MacPhee 1999).
The authors seem to have missed the point
that predation by Homo sapiens has not led to extinctions in the
continent where humans evolved. However, whenever humans migrated to new
continents and islands, they were able to extirpate large animals that
had not evolved either fear of or defences to humans. The same happened
with the introduction of alien predators like rats, cats and stoats to
remote islands.
Top-down regulation is a myth since populations
interact on the basis of economy or effort. See our explanation of the
economics
of exploitation in the
Resource Management
chapter.
The authors continue with a list of examples of trophic
cascades and research done on this.
Trophic cascades are defined as predatory interactions involving three
or more trophic levels, whereby primary carnivores indirectly increase
plant abundance by suppressing herbivores (Menge 1995). . . (more references)
. . However, good empirical examples supporting the existence of such trophic
effects are generally lacking (Scheibling 1996). . . . (The authors mention
the Californian sea otter - urchin - kelp story. Which
may well be flawed like the research done here in NZ) . . In some
areas where sea otters do not occur, fish and lobsters have been implicated
as important predators of urchins. (more doubt) While
the destruction of kelp beds by sea urchins in these areas has been linked
to overfishing of both lobsters and fish, the existence of a direct causal
linkage has received much debate (Scheibling 1996).
. . . . . (more references) . . However there has not yet been any
decline in the extent of urchin barrens in these protected areas (Mediterranean
Sea). Other factors such as recruitment, pollution, disease,
large-scale oceanographic events, urchin harvesting, food subsidies
and availability of shelters may also be important in controlling algal
assemblage structures (Sala et al 1998).
This paper deliberatley fails to discuss
the most important effects on urchins and kelp, those of recruitment failure,
massive kills, pollution and storms, all of which have had a major influence
during and before the experiments. Note how other studies could not replicate
the decline of urchin barrens in protected areas.
The aim of this study was to demonstrate the indirect effects of fishing
on lower trophic levels by experimentally examining the top-down role of
predators in explaining the habitat change documented in marine reserves
in northeastern New Zealand. This was done by:
An urchin-tethering experiment to test whether relative predation levels
on urchins were higher at marine reserve sites and to determine the sources
of predation.
Comparing the density and demography of urchins in the urchin barrens habitat
at multiple sites in two reserves and two non-reserve areas.
Experimental removal of urchins to test whether the observed habitat changes
in the Leigh reserve were consistent with a reduction in urchin populations.
Comparing the distribution of macroalgal communities among reef habitats
between reserve and non-reserve areas.
It is deplorable that this study was motivated
politically, to show that marine reserves work, rather than searching for
more understanding of urchin and kelp populations. The authors chose
the only two marine reserves in New Zealand (out of 15 others) where urchin
habitat disappeared!! Is this by chance?
Materials and methods Study area: see map.
Predation . . . The tethering technique involved inserting a hypodermic needle
(1.2x38mm) through the dorsal and ventral surface of the urchin test, as
far away from the oral-aboral axis as possible. Nylon monofilament was
then threaded through the needle and tied off. laboratory trials found
100% survival of 80 tethred urchins [ranging from 25 to 75mm test diameter
(TD)] after 10 days. Trials in the field found that tether-related mortality
could be reduced by holding the urchins in the laboratory for a week prior
to experimentation. This procedure also allowed the urchins to heal, minimising
the potential effects of coelomic fluid leakage on predation (McClanahan
& Muthiga, 1989) . . .
Daily monitoring enabled detection and replacement of urchins that appeared
to be dying as a result of tethering. In each experiment only four out
of a total of 180 tethered urchins died as a result of tethering. . . .
Fig 1. Location of study sites in the Cape Rodney to
Okakari Point Marine Reserve (CROP) and Tawharanui Marine Park. Circles
indicate sites where the predation experiment was carried out. Inset shows
general location of study area in New Zealand's North island.
This is a well thought-out experiment
with adequate precautions in the laboratory. When urchins bleed (coelomic
fluid leakage) they attract scavengers of which there are many, and it
cannot be guaranteed that the urchins had indeed healed completely. To
do an experiment where this does not play a role, requires careful execution.
However, the two coasts selected are incomparable. Both reserves are facing
north, being much more exposed than the SE facing coast. Three locations
to the west of both reserves are located in shallow water where urchin
barren zones normally don't occur. Results could be interpreted as "north-facing
coasts favour survival of tethered urchins". coscinasterias being
the main predator in the non-reserve group, bears this out. Worse still,
was the north facing slope of Kawau (SE of Tawharanui) ignored as
control site because it produced inconsistent data in other experiments?
Predation on urchins was significantly
higher at reserve sites than at non-reserve sites (F=9.44, P=0.0133), with
the relative odds of predation being 6.9 times higher at reserve sites
(Fig.2, Table 1).
The fate of all small urchins preyed upon at reserve
sites was unknown as the tests were completely removed from their tethers
(Table 2). This could have been due either to predation by fish, which
completely engulf the urchin, or by lobsters breaking up or removing small
urchins. (or by wave action?) At reserve sites
approximately 45% of preyed individuals in the larger size classes showed
patterns of damage characteristic of spiny lobster predation. No urchins
showed signs of spiny lobster predation at non-reserve sites. In most cases
mortality at non-reserve sites could be attributed to starfish (Coscinasterias
muricata) or the gastropod Charonia lampax, both of which are
slow-moving predators. (but which remain on-site,
having an acute sense of smell for stressed animals)
Reader please note that a sleight
of hand is presented here, where the researchers talk about odds of predation,
which is the inverse of survival. The curves on right show survival
rates, with non-reserve sites doing between 30-90% of reserve sites. Leigh
approx 30,50,70% and Tawharanui 40, 80, 80%. Note also that mortality is
critical in the first few days, after which it becomes negligible.
The authors do not attempt to explain this effect.
Table 1
Size class
Number preyed
Unknown%
Lobster%
Coscinasterias%
Charonia%
Reserve
35mm
55mm
75mm
40
23
13
100
56.5
46.2
0.0
43.3
46.2
0.0
0.0
7.7
0.0
0.0
0.0
Non-Reserve
35mm
55mm
75mm
11
7
4
54.5
42.9
0.0
0.0
0.0
0.0
45.5
42.9
50.0
0.0
14.3
50.0
Fig 2. Survival of tethered urchins at reserve (black
circles) and non-reserve (open circles) areas. The mean number of tethered
urchins surviving in each of the three size classes is given for Leigh
and Tawharanui.
We wonder why this experiment was not
continued for days 15, 20 and 30, which would have revealed more interesting
information. Reader please not that the difference between 98% and 96%
survival rate is negligible while this corresponds to mortality rates of
2% and 4%, representing a major difference. Note how survival rates eventually
level off.
Urchin density and size structure Densities of Evechinus in the urchin-grazed habitat varied widely between
sites but were generally lower at reserve sites for both areas (Fig 3).
the density of exposed urchins (Fig 3A) was significantly lower at marine
reserve sites (Tables 1,3 [left out]). Exposed
urchins were 1.7 times more abundant overall at non-reserve sites (Table
1 [left out]). There was no difference in
urchin density between Leigh and Tawharanui areas but there was a significan
interaction between Area and Status (parameters of
the statistical mixer).
Fig 3. Mean density of A exposed and B cryptic urchins,
and C mean size of exposed urchins
from quadrat sampling (n=10) at all reserve (open circles)
and non-reserve sites (filled circles).
This graph shows fewer large urchins
in reserves but a comparable number of recruits. Also those inside reserves
are larger, although not so for Tawharanui. The difference is more pronounced
for Goat Island and almost negligible for Tawharanui. when expressed in
grazing capacity, the difference is likely to become even less significant.
This difference comes to expression in the bimodal size distribution of
urchins inside the Leigh reserve.
Comparisons of urchin populations Populations were more bimodal (having two peaks)
at reserve sites, with very low numbers of urchins between 30 and 50mm
and they generally remained cryptic (hidden in cracks
and under boulders) to a greater size. This pattern was stronger
in the Leigh marine reserve.
The authors make no effort explaining the bi-modality,
which
could well be a symptom of stress, degradation, mortality or recruitment
failure, all three symptoms have been observed in the reserve areas.
The fact that urchins remained cryptic to a greater size may well be related
to the difference in wave exposure between the reserve and non-reserve
sites. It urges caution in interpreting the tethered urchin experiments
where the small urchins (35mm) and arguably also the 55mm ones were indeed
still cryptic for the north facing coasts. If these are omitted from the
experiment, only very little difference in survival rate remains to explain
the disappearance of the barren zones.
Fig 4. Size frequency distribution of all Evechinus
measured during quadrat sampling at each area. Shaded bars indicate poportion
of cryptic urchins.
Urchin removal At the commencement of the experiment in January 1998 Evechinus
densities did not vary between treatments. Densities of urchins ranged
from 1.2 to 2.4 per 0.25m2. Crustose coralline algae (Lithothamnion
and Lithophyllum spp.) were dominant, covering 63-99% of the substratum.
Articulated coralline turf was the other dominant encrusting form with
cover ranging between 0 and 35%. There was no significant difference between
either crustose coralline algae or coralline turf between treatments or
between plots within treatments. Macroalgae were rare at the start of the
eperiment, with Carpophyllum flexuosum which is relatively resilient to
urchin grazing (Cole and Haggit 2001), being the only conspicuous large
seaweed. . . .
After 1 year the control plots remained as urchin barrens dominated
by crustose coralline algae, while the urchin-removal plots had become
dominated by coralline turf, with a mixture of large and small brown algae.
. .
This experiment was conducted in the SE facing
shore near Mathesons Bay, which is characterised by medium exposure and
rock fissures creating ideal isolation for experimental plots. Although
the experiment removed the urchin as grazer, other grazers such as Cookia
sulcata and many fish species still had access.
The experiment shows that large seaweeds will
not necessarily take over, even after one year of isolation. But the small
seaweed species took over (note the difference of an order of magnitude
in the scale of graph D!
This result provides a strong argument against
the kelp win, urchin lose hypothesis! It also provides a strong argument
against high kelp productivity since most productivity by far arose from
small brown algae.
Fig 7 A-D. Response of macroalgae to urchin removal.
The mean abundance of A Carpophyllum flexuosum, B Ecklonia radiata,
C Halopteris virgata and D small brown algae in both control (open
circles) and urchin-removal plots (full circles) following the commencement
of the experiment in January 1998.
Distribution of urchins habitat The general distribution of habitats (Fig. 8) at non-reserve sites
is typical of northeastern New Zealand with shallow fucoid (bladder
weed) assemblages, deep kelp forests (Ecklonia radiata) and
intermediate depths dominated by urchin barrens (Choat & Schiel 1982).
However, at marine reserve sites all depth ranges were dominated by macroalgal
habitats. The proportion of urchin barrens habitat was significantly lower
than at non-reserve sites (Tables 1,7) [7 left out]). The relative odds
ratio for the proportion of urchin barrens at reserve vs non-reserve sites
was 0.2 to 1 (Table 1) or inversly, 5.9 times higher at non-reserve sites.
This pattern was consistent between both areas but varied significantly
with depth (Table 7). Differences . . were greatest in the shallow depth
strata (0-3m and 4-6m) where urchin barrens covered approximately 54% of
available reef at non-reserve sites and only about 20% of the reef at reserve
sites. The deeper strata (7-9m and 10-12m) at both reserve and non-reserve
sites were dominated by macroalgal habitat, predominantly Ecklonia
forest.
Fig 8. Mean percent cover of major habitat types (shallow
fucoids, urchin barrens, turfing algae
and kelp forest), within each depth range for reserve
and non-reserve sites at Leigh and Tawharanui.
The top two bar charts show that the
fucoid zone in Leigh is deeper than elsewhere, wich demonstrates what we
alluded to earlier, that the Goat Island coast is more exposed. Note
how the urchin barrens have been invaded by the kelp forest, which is easy
to see under water. Had Kawau and/or Little Barrier Island been included,
they too would have shown a kelp takeover. Sadly, this has not been done.
Discussion
New Zealand's two oldest marine reserves at Leigh
and Tawharanui support higher predator densities than similar unprotected
areas. Snapper 5.8-8.7 times; spiny lobster 1.6-3.7 times. True,
but this is evident already after five years of no-take protection (see
paper
on this page). The high snapper density is caused by using the baited video
camera method. The low spiny lobster count was caused by the low numbers
in the Tawharanui reserve!
Relative rates of predation on (tethered)
sea urchins were found to be higher in the reserves. True
but by far insufficient to explain the kelp takeover.
The densities of actively grazing urchins (excluding
cryptic urchins) is lower in the reserves.
True
but so they are at Kawau, Little Barrier, Great Barrier and further.
The cover of macroalgal forests is higher in the
reserves. True but so they are at Kawau, Little
Barrier, Great Barrier and further.
These patterns . . . provide experimental evidence
of top-down impact of predators on subtidal reef communities. This
is a sleight of words. Every predator causes top-down impact. However,
the
results do not show sufficient impact to substantiate top-down control,
which is an entirely different matter. It shows a shift in the economy
of exploitation - perhaps. The results are entirely a consequencce of the
1993 kelp death degradation event.
The spiny lobster Jasus edwardsii was found
to be an important predator of sea urchins at marine reserve sites. True.
However, the control sites were ill-chosen. The prickly star Coscinasterias
is not common on exposed coasts.
This is noteworthy, considering that spiny lobsters
were not previously thought to forage in the urchin barrens habitat (Andrew
& Choat 1982, Andrew & MacDiarmid 1991). From
personal observations and those of a professional urchin diver, spiny lobsters
are attracted to animals in distress, wherever they are located.
More importantly, when excluding the small
urchins, the difference in survival rates between reserve and non-reserve
are not enough to base far-reaching conclusions on.Previous experimental studies carried out in the
Leigh reserve acknowledged that both snapper (Andrew & Choat 1982)
and spiny lobsters (andrew & MacDiarmid 1991) were important predators
of Evechinus in northeastern New Zealand, but concluded that predation
by these species was not of sufficient magnitude to substantially alter
urchin populations and cause community-level effects.We
concur. This study does nothing to disprove the two earlier studies.
Their study was carried out after only 4 years
of marine reserve protection. If it were to be repeated now after 25 years
of protection and recovery of predator populations, a larger effect may
be expected. This myth lies at the heart of
so much research done in recent years, blinding researchers from seeing
the obvious. Where is the proof?
This study demonstrates the value of marine reserves
as experimental tools to test ecosystem-level hypotheses at ecologically
relevant scales, previously unfeasible using traditional caging and manipulation
experiments. (Andrew & MacDiarmid 1991). Marine reserves have enabled
us to measure the top-down role of predators in structuring subtidal reef
communities in northeastern New Zealand, as well as the indirect effects
of fishing on the trophic structure of reef communities.
This
study does not give enough evidence for the above statement. When other
areas are taken into account (Kawau, Little Barrier, Great Barrier, Rakitu,
Mokohinau) it can be proved that this study is a failed experiment.The
map shows how the authors have observed kelp invasions inside two marine
reserves (red areas), but failed to observe similar effects elsewhere,
particularly in the yellow areas nearby the study area (yellow areas).
All areas where we observed kelp invasion of the urchin barrens were located
where previously kelpbed death has been observed. This can still be confirmed
today (2006).
It is unclear whether, or to what extent, these findings
can be extrapolated to other regions where urchin barrens are less common
and interactions between trophic levels are weaker. They
CANNOT.
Reader please note that in 2007 most of the urchin
barren zones along the entire NE coast of the North Island had either disappeared
or become infested with seaweeds. It has nothing to do with marine protection
or fishing.
Protection of exploited fish in temperate
regions: high density and biomass of snapper Pagrus auratus
(Sparidae) in northern New Zealand marine reserves.
Trevor J Willis, Russel B Millar, Russ C Babcock.
Journal of Applied Ecology (2003) 40,
Reader
please note that scientist Trevor Willis has taken the unusual step to
have this rebuttal removed, by complaining to the publisher about us 'infringing
both his intellectual and moral rights'. The publishers then quite wrongly
threatened Seafriends and Dr Floor Anthoni in a heavy-handed way. As you
can see, our valid review and criticism has hit a raw nerve for which these
scientists have no reply. We urge you to read the exchange of letters between
us and the publisher, which will forever mark a black day for science.
Also read other attacks on our integrity in say/opinion.htm
The use of marine reserves as tools either for conservation or fisheries
management requires rigorous empirical evidence for the recovery of exploited
species within them. True. It also requires rigorous
empirical evidence for sustainability, i.e. not losing non-fished species.
Little work has been done on this, but in the meantime (2009) evidence
has accrued. See shellfish
collapse in NZ., fish decline at the
Poor Knights and monitoring results
at Goat Island.
The relative density and size structure of snapper
Pagrus
auratus (Sparidae), an intensively exploited reef fish species, were
measured using a baited underwater video, inside and outside three northern
New Zealand marine reserves (Leigh marine reserve, Hahei marine reserve
and Tawharanui marine park) every 6 months from October 1997 to April 1999.
Log-linear modelling showed that relative total density
and egg production of snapper were higher in three reserves than in fished
areas. Snapper that were larger than the legal minimum size were estimated
to be 14 times denser in protected areas than in fished areas, and
their relative egg production was estimated to be 18 times higher.
In the Leigh reserve, legal-size snapper were larger than legal-size snapper
in fished areas. This would imply that the
snapper stocks outside have been fished down to below 5% of pristine levels,
which is not borne out by fisheries statistics. The Leigh marine reserve
data swamps all other because it is a special place, incomparable with
other places in NZ (see recurring themes
above).
At the Leigh reserve, snapper density consistently
peaked at the reserve centre and declined towards either boundary, which
suggests that snapper became increasingly vulnerable towards the reserve
boundaries. However, the two other reserves
disprove this statement. The authors forget to mention that the centre
of the Leigh reserve offers the most shelter, is nearest to where people
feed and where activity is and juts out furthest in sea offering much
clearer water than elsewhere.
Inshore snapper density was significantly higher
in autumn than in spring, supporting previous suggestions that snapper
make regular onshore-offshore migrations that might be related to spawning.
We
suggest that the observed recovery of snapper populations within reserves
is attributable to immigration of individuals from fished areas that take
up residency within reserves, rather than juvenile recruitment.
This
study does not provide the evidence that any of the measured snapper stay
resident. Seasonal fluctuations suggest that up to 80% of snapper move
in and out of the reserve, thus escaping effective protection. The suggestion
is important for reserve design and the expectations people have of marine
reserves. Most snapper are NOT protected by marine reserves. The three
marine reserves are hot spots and not representative of the coast.
Synthesis and applications. This study demonstrates
the effectiveness of marine reserves for protecting an exploited species
previously thought to be too mobile to respond to area-based protection.
We suggest that the protection of fish populations within reserves might
slow reductions in genetic diversity caused by size-selective mortality
brought about by exploitation. This study
shows exactly the opposite. The reasons for snapper to be more numerous
inside a reserve in summer, must be explained differently.
Introduction It has been widely suggested that marine reserves (areas of sea permanently
closed to all forms of fishing or disturbance),
in addition to performing a conservation function, might be of long-term
benefit to fished stocks. The potential benefits are many [oops,
see marine conservation and FAQs]
and have been described extensively (Roberts & Polunin 1991; Allison,
Lubchenco & Carr 1998; Bohnsack 1998; Jennings 2000).
In essence, the ideal is protection of a portion of an exploited stock,
with the expectation that the biomass of targeted species within protected
areas will rebuild to approach unfished densities. Density-dependent processes
might then cause emigration of adults from the source (reserve) population
to fished areas, either by passive diffusion (Beverton & Holt 1957)
or by displacement of individuals caused by space limitation (Kramer &
Chapman 1999). Additionally, spawning activity within the reserve by greater
numbers of large individuals should result in greater production of gametes
than in similar, unprotected areas.
. . . (the authors discuss computer models in studies) . . . While
these effects are generally positive, on a case-by-case basis evidence
for substantial recovery in populations is often limited (Jones, Cole
& Battershill 1993; Rowley 1994; but see Russ & Alcala 1996a; Wantiez,
Thollot & Kulbicki 1997; Edgar & Barrett 1999). There are three
main reasons.
First, exploitation in any geographical region tends to begin with large
predatory species (Pauly et al. 1998) that are less common than species
at lower trophic levels and therefore more difficult to monitor with sufficient
statistical power (Cole, Ayling & Creese 1990; Paddack & Estes
2000). Moreover, larger predators tend to be slower to grow and reproduce,
which means that population-level responses may be slow (Jennings, Reynolds
& Mills 1998).
Secondly, with few exceptions (Edgar & Barrett 1997, 1999; Wantiez,
Thollot & Kulbicki 1997), the design of marine reserve surveys has
often been spatially or temporally confounded (or both) so that the results
must be interpreted cautiously.
Finally, field methods used to assess fish density have sometimes been
subject to biases caused by intra- or interspecific behavioural plasticity
. . . . .
. . . . In this study we examined the effects of marine reserve protection
on the density and size of snapper at three coastal marine reserves of
varying age in north-eastern New Zealand. The aim of the study was to assess
the general effects of reserves by using spatially and temporally replicated
surveys. Specifically, we wished to
determine the magnitude of differences in snapper density and size between
reserve and adjacent fished areas, and
quantify seasonal and interannual variability in snapper density and size.
Methods Study areas The three reserves were the Cape Rodney to Okakari Point (Leigh) Marine
Reserve, Tawharanui Marine Park and Te Whanganui a Hei (Hahei) Marine Reserve
( Table 1 and Fig. 1a). All three are complete no-take areas administered
under different legislation and by separate government departments (the
reserves are controlled by the Marine Reserves Act 1971 and administered
by the Department of Conservation, whereas the park is controlled by
the Fisheries Act 1983 and administered by regional authorities). The management
of the Tawharanui Marine Park has no nearby local community that can provide
day-to-day surveillance to assist enforcement because the adjacent land
is a regional park.
Incorrect. Local communities have not been involved
in managing any of the three reserves. This is done centrally by government
departments.
Fig. 1. (a) Map of the Hauraki Gulf and environs, showing
the location of the three reserves surveyed in this study. Depth contours
are in metres (?). The inset shows the location
of the area in the North Island of New Zealand. (b) Details of the three
reserves surveyed, showing reserve boundaries (dashed lines) and survey
areas. Top: Leigh (1977); centre, Tawharanui
(1982);
bottom, Hahei (1992).
At each sampling site, observations of snapper relative density were
made using a baited underwater video (BUV) system (Willis & Babcock
2000). This system was developed in response to difficulties in accurately
sampling a species whose behavioural reactions to divers vary markedly
between sites (Cole 1994; Willis, Millar & Babcock 2000). Fish feeding
by visitors to the Leigh marine reserve has resulted in snapper exhibiting
diver-positive behaviour at some sites, whereas elsewhere they are wary
of divers, and outside the reserve they actively avoid divers. Use of a
remotely deployed sampling method eliminates this source of bias.
We have serious doubt about the accuracy
of the BUV system as explained in Recurring
Mistakes. Just the way snapper develop diver-positive behaviour, they
also develop BUV-positive behaviour with pilchards as reward.
The BUV system consisted of a Sony XC-999P
high-resolution colour camera mounted on a stainless steel stand 115 cm
above the substratum and faced straight down. A bait holder (containing
c. 200 g of pilchard
Sardinops neopilchardus Steindachner) was attached
to the triangular base of the stand so that it lay in the centre of the
cameras field of view. The base was marked with cable ties, and the distance
between them was measured to allow spatial calibration of digitized images.
This allowed accurate estimation of the lengths of fish responding to the
bait (Willis & Babcock 2000; Willis, Millar & Babcock 2000).
Replicate deployments (n = 4 per survey area at Leigh
and Tawharanui, and n = 5 per area at Hahei) were made on soft substrata,
either immediately adjacent to or within 50 m of the reef. The BUV assembly
was lowered to the sea floor from an anchored vessel, and deployed for
30 min from the time contact was made with the bottom. At the laboratory,
video footage was analysed (frame-by-frame where necessary) to determine
the maximum number of snapper (MAXsna) in the field of view during each
30-min sequence. Individual fish lengths (FL) were measured from calibrated
images using the Mocha® image analysis software (Jandel Corporation).
Measurement error using this method was typically < 20 mm (Willis &
Babcock 2000). Fish were generally only measured from images taken at the
time MAXsna was recorded. On a few occasions fish that occurred elsewhere
in the sequence were measured because they were obviously different fish,
by virtue of size (i.e. differed from MAXsna measurements by >100 mm).
Small snapper that appeared early in the sequence were the most frequent
additions to the data set, but sometimes one or two large fish were measured
in this way. Although this meant that some fish were not measured, it also
avoided repeated measurement of the same individuals.
The entire method is aimed at attaining
the maximum result.
. . . (the authors explain how biomass and egg production were estimated,
and explain the statistical analyses applied)
The formula for daily batch fecundity
(F) of Zeldis & Francis 1998 is F = 73.9 x W -7793 where W =
snapper biomass or weight. For ease of reference, it approximates to F=
(W - 105) x 74, where snapper need to weigh over 105g to produce 74 wet-weighted
eggs per gram of body weight in each spawning batch. The formula was derived
from snappers weighing between 200g and 5kg (65cm).
Results EFFECTS OF MARINE RESERVE PROTECTION ON P. AURATUS DENSITY, BIOMASS
AND EGG PRODUCTION
Biomass per BUV deployment and density of legal-size snapper (LEGsna)
were higher in the reserve than adjacent non-reserve areas at all three
locations and for all four surveys (Figs 2 and 3). In particular, the Leigh
reserve recorded the highest value of LEGsna on all four survey occasions,
and the highest density of snapper of all sizes (MAXsna) on all but the
April 1999 survey, when the Hahei reserve MAXsna was boosted by large numbers
of sublegal fish.
The graphs show the number of snapper (above)
and their relative biomass (below) for the two years of observation. It
shows that the number of snapper in each reserve were comparable but that
the ones in Leigh were much larger. It also shows that the numbers and
sizes of snapper outside the reserves were similar for all areas. The main
conclusion could be either that Leigh is a special area attracting large
fish, or that Leigh has larger fish because it is older. For each possibility,
proof is still lacking.
It is regrettable that the authors did only two
surveys per year, rather than one for each of the four seasons. Because
of this, it is not known when maximum and minimum densities occur.
Fig 2b shows that the biomass inside two marine
reserves is 2-3 times that outside, with Leigh being an exception.
Fig. 2. Mean reserve (filled symbols) and non-reserve
(open symbols) snapper Pagrus auratus relative density at Leigh,
Hahei and Tawharanui from November 1997 to April 1999. (a) Total numerical
relative density, all size classes (MAXsna); (b) relative biomass.
Fig. 3. Mean reserve (filled symbols) and non-reserve
(open symbols) snapper Pagrus auratus numerical relative density
at Leigh, Hahei and Tawharanui from November 1997 to April 1999. (a)
Fish > minimum legal size (LEGsna); (b) fish < minimum legal
size (JUVsna).
Fig. 4. Predicted mean LEGsna by survey area at Leigh,
Hahei and Tawharanui. Vertical dashed lines represent reserve boundaries,
solid symbols are reserve (R) areas and open symbols are non-reserve (NR)
areas.
Fig. 5. Predicted mean JUVsna by survey area at Leigh,
Hahei and Tawharanui. Vertical dashed lines represent reserve boundaries,
solid symbols are reserve areas and open symbols are non-reserve areas.
The above graph (Fig3) shows that legal-sized
snapper are more abundant inside reserves during the end of summer, but
Leigh has more legal sized-snapper all year round. Juveniles do not make
a distinction between reserve or non-reserve.
Fig 4 and 5 show how adult and juvenile snapper
are distributed along the reserve's coast. As expected, juveniles are not
affected by marine reserves but adults do. Only in Leigh does the density
increase towards its centre. It again demonstrates that Goat Island is
a special place which cannot be compared to others. It is remarkable
that the number of juveniles increases in areas 9-13 outside the Leigh
reserve. But this coast is also more sheltered and influenced by currents.
It is important to note that the differences
between a reserve of 7 years old (Hahei, 1992) and one of 17 years (Tawharanui,
1982) is negligible. Only Leigh (1977, 22 years old) stands out, but it
is an oasis with an island, next to a long beach.
Discussion Surveys of three marine reserves in northern New Zealand, repeated
biannually for 2 years, have allowed estimation of the effects of protection
on snapper populations, as well as location and seasonal effects. The design
of the study reduced the risk of location-specific biases that may have
been present due to the lack of before data, which in marine reserve
studies are often unobtainable. The data presented here demonstrate large
differences in relative density of the heavily exploited sparid fish P.
auratus between marine reserves and adjacent fished areas in northern
New Zealand. Log-linear modelling indicated a common status (reserve or
non-reserve) effect corresponding to a 14-fold increase of legal-size
snapper in the reserve compared with the adjacent non-reserve areas,
despite significant between-location and between-survey variability in
densities.
Repeatedly, all studies where the
Leigh reserve is included, show that Leigh is an exception to the rule.
When do scientists become honest by excluding Leigh from their data, certainly
when such data is obtained by BUV. Fig 2b shows clearly that there are
no large differences (about 2 times by BUV standards) between reserves
that are typical of our coast and non-reserve sites. This is also more
in line with studies done overseas. Also in winter/spring, the differences
disappear.
. . . . (more statistical niceties) . . .
At all three reserves, counts were much lower in the spring (OctoberNovember)
surveys than in the autumn (AprilMay) surveys. This pattern agrees with
previously recorded observations of seasonal increases in snapper density
on reefs and soft sediment bottoms. Trawl surveys have indicated that the
abundance of snapper at inshore Hauraki Gulf locations fluctuates seasonally
(Paul 1976). Within the Leigh reserve, high densities of juvenile fish
belonging to the 0+ and 1+ year classes have been described on reef habitats
during spring and summer (DecemberMarch) but densities were very low in
winter (Kingett & Choat 1981). Similarly, Francis (1995) suggested
that observed seasonal changes in juvenile snapper density on soft sediment
bottoms might be attributable to movement of fish onto reefs in early summer.
Similar evidence for seasonal changes in snapper abundance has been recorded
from Japan (Matsumiya, Endo & Azeta 1980; Kiso 1985).
Interestingly, Kingett & Choat (1981) did not detect the seasonal
fluctuations in the density of older fish that were found in this study,
possibly due to bias caused by the presence of diver-habituated resident
fish (Cole 1994; Willis, Millar & Babcock 2000). Similarly, the angling
experiment of Millar & Willis (1999) did not detect seasonal variability
(June vs. December) in snapper catch-per-unit-effort data at Leigh. This
could be due to capture biases, but it is also likely that June and December
are both part way through the emigration and immigration (respectively)
of snapper to inshore reefs.
It is regrettable that the experiment
does not show four sample points for each year. Yet the need for this could
have been foreseen.
The large seasonal fluctuations in snapper density have implications for
marine reserve monitoring and the prediction of potential reserve benefits
to fisheries. First, there is need for standardization of the timing of
surveys to determine reserve effects. If different reserves are surveyed
at different times of year, the results will not be comparable, and will
give misleading impressions of the relative effectiveness of the different
reserves.
This has indeed happened in several surveys.
This may apply to species other than snapper. Theoretical reviews have
predicted that migratory species, or species with moderate mobility, will
not benefit significantly from marine reserve protection (Kramer &
Chapman 1999). In this case, however, the density of a migratory species
is much higher within reserves than in fished areas. It appears that most
snapper are seasonally mobile, but some individuals have shown a marked
degree of site fidelity (Willis, Parsons & Babcock 2001). Thus, generalizations
about the entire species are inappropriate, and theoretical predictions
made from such generalizations are likely to lead to incorrect conclusions.
. . . . .
As suggested by Cole (1994), high abundance of snapper at the reserve
centre might be a response to (i) differences in habitat quality, (ii)
hand feeding of fish by the public or (iii) higher levels of surveillance
at the reserve centre. It is also possible that these factors are of secondary
importance to the relative vulnerability of site-attached fish (Willis,
Parsons & Babcock 2001) to fishing pressure at the reserve boundaries.
Goat Island is a special place with a
mix of desirable habitats, exposure and shelter, extensive shellfish feeding
grounds and sleeping habitat, all of which are important to snapper, particularly
large ones. The outside of Goat Island, in the centre also has the clearest
water.
. . . . . . There are two potential direct benefits of marine reserves
to fisheries: (i) enhancement of spawning stock biomass and (ii) spillover
of adults to enhance local fisheries (Roberts & Polunin 1991; Rowley
1994; Allison, Lubchenco & Carr 1998; Bohnsack 1998; Horwood, Nichols
& Milligan 1998). Seasonal peaks in inshore snapper density coincide
with the spawning season and post-spawning period (Crossland 1977; Scott
& Pankhurst 1992), implying that marine reserves protect both resident
fish and some proportion of migratory fish during spawning, assuming that
they spawn within reserves. However, greater output of eggs need not necessarily
translate into production (Francis 1993).
Variability in larval mortality means it has proven difficult to
determine any relationship between spawner abundance and recruitment (Myers
& Barrowman 1996; Gilbert 1997), and increased contributions from
reserves are likely to become important only when stocks are overfished
to low levels. If reserves are to have measurable effects, they may have
to be large, perhaps to the detriment of the fishery (Parrish 1999). However,
the inability to detect a measurable effect on production or recruitment
does not mean some beneficial contribution is not being made (Lauck et
al. 1998). For example, our data suggest that a reserve the size of
Leigh (c. 5 km of coastline) might conservatively produce a quantity of
snapper eggs equivalent to that produced by c. 90 km of unprotected coastline.
This is not borne out by this experiment.
The claim of 14 times is caused by the Leigh data set swamping the two
other data sets. By all scientific standards, it should have been removed
as a spurious dataset biasing the experiment unduly. The remaining bias
comes from the BUV counting method, whose linearity has not been proved.
It is a method that exaggerates.
The term spillover implies that density-dependence (whether via resource
limitation or territoriality) actively displaces fish across reserve boundaries,
where they become available to the fishery (Kramer & Chapman 1999).
In this study, the survey areas adjacent to the reserve boundaries generally
contained the lowest overall density of P. auratus of non-reserve
areas. This was most likely because of concentrated recreational fishing
effort at those locations (especially area 9 at Leigh; authors personal
observation) caused by perceptions that catch rates next to the reserve
are likely to be high. If emigration from the reserve has been occurring,
numbers were too low to be detected with the current methodology. In this
regard, the importance of knowing the distribution of fishing effort outside
reserves cannot be understated.
The outside areas are all different habitats.
To the west under influence of the Pakiri Beach and to the SE they offer
more shelter. They cannot really be compared with Goat Island.
This research was conducted with a measuring
device (BUV) which fails the scientific requirements for measuring devices
and cannot be used for quantitive values. The data obtained from Leigh
completely swamps all other data such that the conclusions in their generality
are not substantiated. But it is clear that marine reserves do have more
snapper and crayfish.
Effects of marine protection
on benthic reef communities: northeastern New Zealand Nick T Shears, Russell C Babcock, 2001
Report to the Department of Conservation. 31p.
[even after repeated requests, DoC refused to
give us a copy of this report,
so here is the authors' executive summary obtained
from the Internet]
Executive summary Ecological theory predicts that when the density of predators changes,
there may be reactions in the form of changes in density at lower trophic
levels. In the context of the marine communities of rocky reefs in northeastern
New Zealand this means that where marine reserves allow predator densities
to increase, the densities of their herbivore prey (e.g. grazing urchins)
should decrease, and that of macroalgal primary producers increase.
The same ecological theory demands that
such changes happen as gradually as large predators establish themselves.
The observed disappearance of urchin barrens however, happened rather quickly
between 1995 and 1998, in one reserve of 24 years of age (Leigh, 1977)
and at the same time also in a younger but nearby one of 19 years (Tawharanui,
1982).
To test the generality of this prediction, subtidal reef communities were
compared between six northeastern New Zealand marine reserves and adjacent
unprotected areas. The marine reserves studied included the Cape Rodney
to Okakari Point (Leigh), Long Bay - Okura, Poor Knights Islands, Tuhua
(Mayor) Island and Whanganui A Hei (Hahei) marine reserves and the Tawharanui
Marine Park. These areas are located across a large geographic and environmental
gradient and varied in size and age of protection. All were completely
no-take and included extensive subtidal reef systems. It has previously
been shown in the reserves at Leigh and Tawharanui that increases in predator
densities, following the cessation of fishing, has resulted in declines
of urchin populations and a subsequent increase in macroalgal habitats.
A cause and effect relationship has never
been demonstrated. See also the research exposed above. But the decline
of urchin populations could readily be observed, as it happened in 1994/95.
After the 1993 kelpbed death, the new kelp invaded the urchin habitat because
(i) urchins wandered into the wide kelp zone, (ii) urchins died from mass
mortalities, (iii) the kelp plants reached maximal growth all at the same
time and (iv) there were no storms to re-create the urchin barrens.
Our findings support this with the extent of urchin barrens habitat and
the overall density of urchins being significantly lower in these reserves
than on adjacent coasts.
However, the other reserves refute these
findings, as do a number of other places around the outer Hauraki Gulf.
The use of the word 'significant' is suspect, since it usually means 'just
detectable'. Why not quantify the difference such as 10%, 2-3 times or
so?
These patterns were not consistent for the other marine reserves examined.
While urchin densities were generally lower at marine reserve sites, the
relative importance of the habitat forming sea urchin, Evechinus chloroticus
and subsequently the extent of urchin barrens habitat, varied considerably
both within and among areas. There were also differences in algal assemblages
among areas and between reserve and non-reserve sites. Only at Long Bay
and Tuhua Island were no differences detected between reserve and non-reserve
sites. This is not surprising for Long Bay, which is very sheltered [and
very dirty and very shallow] and Evechinus does not play
an important structuring role on the reefs. For Tuhua Island the lack of
differences between reserve and non-reserve sites may be due to the reserve
only being 7 years old at the time of sampling.
Tuhua is located in the Bay of Plenty
where the dense plankton blooms of 1991/92 did not happen. However, in
2002 they suffered mass urchin mortality.
The importance of environmental variables (wave exposure, turbidity and
sediment) in explaining the difference in algal communities observed between
reserve and non-reserve sites was tested using multiple regression. The
differences in algal assemblages at Leigh and Tawharanui, between reserve
and non-reserve sites, could not be explained by environmental variables,
thus supporting a significant reserve effect. However, similar differences
observed at Hahei between reserve and non-reserve sites, were confounded
by differences in wave exposure and turbidity between the reserve and unprotected
reference sites.
The control sites chosen around Tawharanui
and Leigh are facing southeast rather than north, or they are located in
shallow water. These places are unsuitable control sites. The Hahei effect
also applies to the control sites around Leigh and Tawharanui. The use
of the 'statistical mixer' method hides whether the Leigh marine reserve
data swamps that of Tawharaunui.
For the Poor Knights Islands, when compared to the Mokohinau Islands as
the reference location, reserve status remained significant even though
there was a significant effect of wave exposure. Given that this reserve
had only been completely no-take for approximately one year, trophic level
effects due to an increase in predators are not likely and the observed
differences were probably due to other environmental differences between
these island groups.
After becoming a fully protected marine
reserve in 1999, the number of snapper suddenly increased spectacularly,
including large snapper but the islands have only a small rocklobster population,
even though these have been protected since 1981. The Mokohinau Islands
have experienced extensive kelpbed death and disappearance of their urchin
barren zones, thus confounding the picture.
While reserve-related differences in benthic communities can only be detected
for the two oldest reserves at this stage, we may expect continued monitoring
to demonstrate similar patterns at other reserves after longer periods
of protection. This study demonstrates the importance of how benthic communities
vary over environmental gradients in order to detect marine reserve effects.
The implementation of pre-reserves sampling programs will help avoid confounding
environmental and spatial effect when assessing the effects of marine reserve
protection.
The main point is that this study does
not prove anything at all. By any standard, it is a failed experiment from
which no conclusions can be drawn. If it takes so much effort to demonstrate
the effects of marine reserves, they are perhaps too small to be of any
practical value. More monitoring and pre-and post-reserve sampling will
not change this.
Indirect effects of marine reserve protection
on New Zealand's rocky coastal marine communities.
Shears NT, Babcock R C (2004):
DOC Science Internal Series DSIS192. (available on the DOC web site)
In a recent study, marine scientists quantified
coastal marine communities in 13 marine reserves spanning the length of
New Zealand. They looked at algal communities and their grazers, particularly
the green sea urchin
Evechinus chloroticus. They also noted environmental
variables such as slope of the substrate (rock) and made an estimate
of the fetch (distance over open water) as a measure of wave exposure.
To their credit, they also included degradation variables of Secchi
disc visibility (the opposite of turbidity) and the percentage
sediment
cover. They also included the maximum transect depth even though
they did not dive deeper than 12m. Unfortunately they did not include
maximum sand depth which is a direct indication of maximum storm damage
and where barrens should occur.
Their most important result is the proof that
degradation
(=turbidity + sediment) is by far the most decisive factor
on what grows where. In other words, the seascape cannot be understood
without understanding degradation. Yet it took us over 15 years of
prodding to get them interested in this very important phenomenon (and
they are still not interested)! Note how the presence of sea urchins also
plays a role (of course) but that
marine reserves have practically no
effect at all (not measurable), proof that they do not have an effect
on urchin barrens and also that they do not save the environment against
degradation. Why did the report not mention these points? These results
also refute the urchin barrens or top-down trophic cascades
theory as espoused by Babcock et al.
Effects of environment
variables 25 turbidity/ visibility
18 sediment cover
10 maximum depth of
transect
7 slope, aspect
6 fetch, wave
exposure
4 sea urchins
in the open
0.5 marine
reserve or not
update As we keep monitoring the environment over as large an area as possible,
we observe changes and document these photographically. This chapter intends
to bring you uptodate with the latest observations. Also overseas studies
are appearing that refute the trophic cascades theory and resulting urchin
barrens.
November 2004 In a CSIRO marine and atmospheric research seminar at the University
of Tasmania, Dr Russ Babcock reports that there is no difference between
fished and non-fished areas: "Studies from north-eastern New Zealand provide
some of the best evidence, so far available, that kelp forest can be converted
to coralline algal dominated barrens as an indirect effect of fishing for
lobsters and fish." None of your or anyone else's
studies has proved this.
"A reversion from urchin barrens to kelp forest has been shown in areas
of north-east NZ where lobster densities were 1.6-4.0 times higher than
in fished areas". This statement has been refuted
and rebutted extensively by us. Why not mention all the failed experiments?
"While such barrens are present in many parts of Australasia and around
the world, their distribution is far from uniform." When
you consider them as storm barrens, their distribution is entirely consistent. "For example, at Rottnest Island in Western Australia,
habitats
are relatively uniform across fished and unfished areas, despite a 6.5
fold difference in lobster density. Such inconsistencies present real
difficulties for predicting marine ecosystem responses to fishing and consequently
to the implementation of ecosystem based fisheries management. Differences
between the two systems are likely to be the result of different ecosystem
dynamics rather than differences in levels of fishing pressure."
Why
not stop believing in the myth and spreading it, Russ? Read about
storm
barrens.
September 2004, July 2005 We visited Niue to study the devastating effect of Tropical Cyclone
Heta (Jan 2004) and found perhaps the deepest storm barrens in the world,
as Niue does not have a continental shelf to diminish the destructive power
of storm waves, and its clear waters support an exceptionally deep photic
zone. A major storm happens here about once in a decade. Calcareous crustose
algae and filamentous algae proved very productive, sustaining schools
of grazing fish by day and huge numbers of sea urchins of many species
by night. We then realised that major storms may also be decisive on the
ecology
of the Kermadec Islands, visited
by us in May 2002.
2006 Ecological Role of Purple Sea Urchins John S. Pearse
Science 10 November 2006: Vol. 314. no. 5801, pp. 940 - 941
Sea urchins are major grazers in shallow seas worldwide. Purple sea
urchins (S. purpuratus) and other strongylocentrotid sea urchins of the
Northern Hemisphere are particularly important and are the most intensively
studied. A delicate balance between sea urchin grazing and kelp forest
productivity leads to stable states that alternate between luxuriant kelp
forests and relatively species-depauperate sea urchin "barrens". Curiously,
the densities of sea urchins are often similar within kelp forests and
sea urchin barrens. Within kelp forests, sea urchins are nearly stationary,
feeding on captured pieces of kelp litter ("drift kelp") that are produced
and shed in high quantities from the kelp plants. However, when the
kelps are removed by storms or El Niño events, the remaining
sea urchins actively forage on young kelp recruits and on drift kelp brought
in from elsewhere, preventing the reestablishment of the kelp forest. The
sea urchins can be decimated by storms or diseases, allowing the kelp forest
to return to the area. Wow, finally someone who has
the right perspective. From here it is but a small step to storm
barrens.
March 2007 Having visited the east coast of New Zealand from the very north to
the Bay of Plenty, the situation is now that urchin barrens have been overrun
by the kelp forest everywhere, whether marine reserve or not. It has happened
in the near-shore environment as well as on remote islands: Cavalli Is,
Poor Knights, Mokohinau, Cuvier, Merury Is, Mayor I.
In all cases we observed an equal presence of dinoflagellate slime
(Ostreopsis)
and absence of major grazers such as Cooks turban Cookia sulcata,
sea urchin Evechinus chloroticus and others. These observations
establish firmly the correlation between slime and the absence of sea urchins,
and also that marine protection has no influence whatsoever. The evidence
is there for all to see and it won't go away!
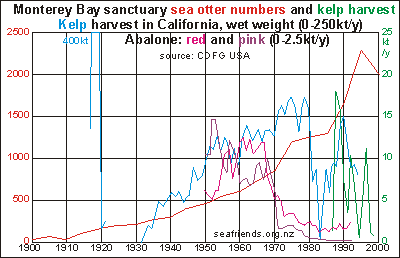 The
Monterey Bay National Marine Sanctuary has been closely monitored for many
years. The graph shows how sea otter numbers (red curve) increased from
near-zero in 1900 to over 2000 in 2000, proof that they had been hunted
to near extinction. However, recently their numbers have been declining
but why is not known. The kelp harvested here is mainly the giant kelp
Macrocystis
pyrifera (and Nereocystis luetkeana) which prefers sheltered
waters, but even so is erratic in its year to year densities. They are
also easily removed by waves. The blue curve shows total kelp harvest in
all of California, climbing as otter numbers increase, suggesting that
there is a linkage between the two. But there are inconvenient exceptions.
Remember that the influences of market demand and price, regulation, nutrient
supplies and degradation can be decisive. The green curve shows kelp harvested
inside the sanctuary, declining as otter numbers are increasing. Kelp is
harvested by cutting the floating fronds off their stipes. The seaweeds
are used to cultivate abalone, and they are exported to Asian countries.
The
Monterey Bay National Marine Sanctuary has been closely monitored for many
years. The graph shows how sea otter numbers (red curve) increased from
near-zero in 1900 to over 2000 in 2000, proof that they had been hunted
to near extinction. However, recently their numbers have been declining
but why is not known. The kelp harvested here is mainly the giant kelp
Macrocystis
pyrifera (and Nereocystis luetkeana) which prefers sheltered
waters, but even so is erratic in its year to year densities. They are
also easily removed by waves. The blue curve shows total kelp harvest in
all of California, climbing as otter numbers increase, suggesting that
there is a linkage between the two. But there are inconvenient exceptions.
Remember that the influences of market demand and price, regulation, nutrient
supplies and degradation can be decisive. The green curve shows kelp harvested
inside the sanctuary, declining as otter numbers are increasing. Kelp is
harvested by cutting the floating fronds off their stipes. The seaweeds
are used to cultivate abalone, and they are exported to Asian countries.
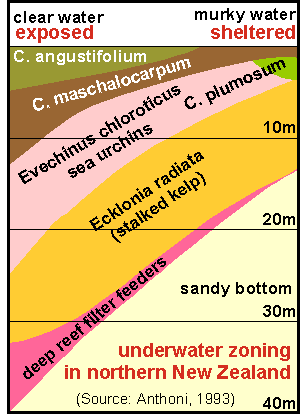
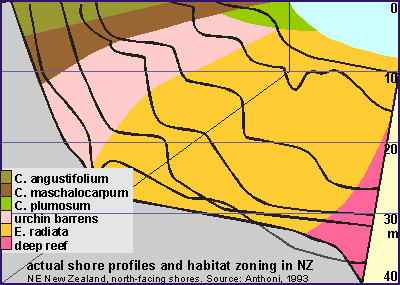
 Because
the plankton blooms between October and December 1992 were exceptionally
dense, a few exploratory dives were made to observe plant life in the dark.
At the peak of plankton blooms, visibility was less than 2m at the surface
and the light at 20m depth was so poor that the sandy bottom could hardly
be distinguished from the dark rock. It was darker than a moon-lit night.
In November the kelp turned dark brown and started to look like wet kraft
paper on woody stalks. By mid December, their crowns fell apart when touched
but in shallower water the kelp appeared entirely unaffected. It was clear
that a major event had happened, and we informed the Marine Laboratory
accordingly. The photo shows what the dead kelp forest looked like.
Because
the plankton blooms between October and December 1992 were exceptionally
dense, a few exploratory dives were made to observe plant life in the dark.
At the peak of plankton blooms, visibility was less than 2m at the surface
and the light at 20m depth was so poor that the sandy bottom could hardly
be distinguished from the dark rock. It was darker than a moon-lit night.
In November the kelp turned dark brown and started to look like wet kraft
paper on woody stalks. By mid December, their crowns fell apart when touched
but in shallower water the kelp appeared entirely unaffected. It was clear
that a major event had happened, and we informed the Marine Laboratory
accordingly. The photo shows what the dead kelp forest looked like.
 In
the first week of January 1993 tropical cyclone Oli struck, and when diving
became possible again, we saw that the whole kelp forest below 15m had
disappeared. In the months following, the kelp died back further to effectively
disappear where once the forest stood. Some shallow water dieback was also
evident but this was caused by the waves of cyclone Oli. The second photo
shows what an extensive sea urchin barren looked like (June 1995, Waterfall
Reef), with a cluster of sea urchins in the foreground and a leatherjacket
(Parika
scaber) above. Note that the kelp had not yet invaded the barrens.
In
the first week of January 1993 tropical cyclone Oli struck, and when diving
became possible again, we saw that the whole kelp forest below 15m had
disappeared. In the months following, the kelp died back further to effectively
disappear where once the forest stood. Some shallow water dieback was also
evident but this was caused by the waves of cyclone Oli. The second photo
shows what an extensive sea urchin barren looked like (June 1995, Waterfall
Reef), with a cluster of sea urchins in the foreground and a leatherjacket
(Parika
scaber) above. Note that the kelp had not yet invaded the barrens.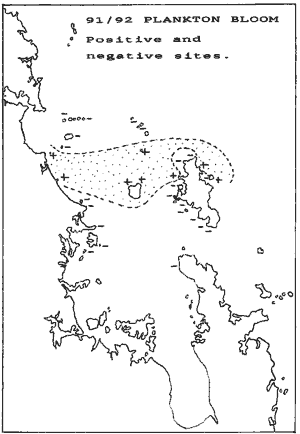
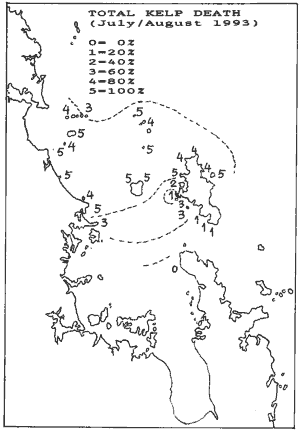
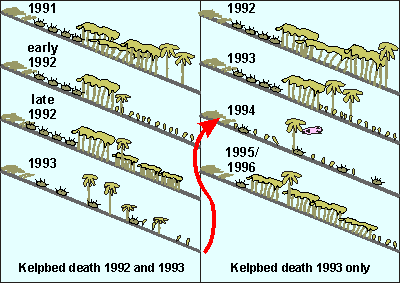 This
diagram depicts our observations in the years preceding and following
the kelpbed death events. On the left the situation where the kelp bed
was removed twice, first partially in 1992, then fully in 1993 (Little
Barrier, Simpsons Rock, Arid Island). On right the situation where the
kelp was removed once, but completely.
This
diagram depicts our observations in the years preceding and following
the kelpbed death events. On the left the situation where the kelp bed
was removed twice, first partially in 1992, then fully in 1993 (Little
Barrier, Simpsons Rock, Arid Island). On right the situation where the
kelp was removed once, but completely.
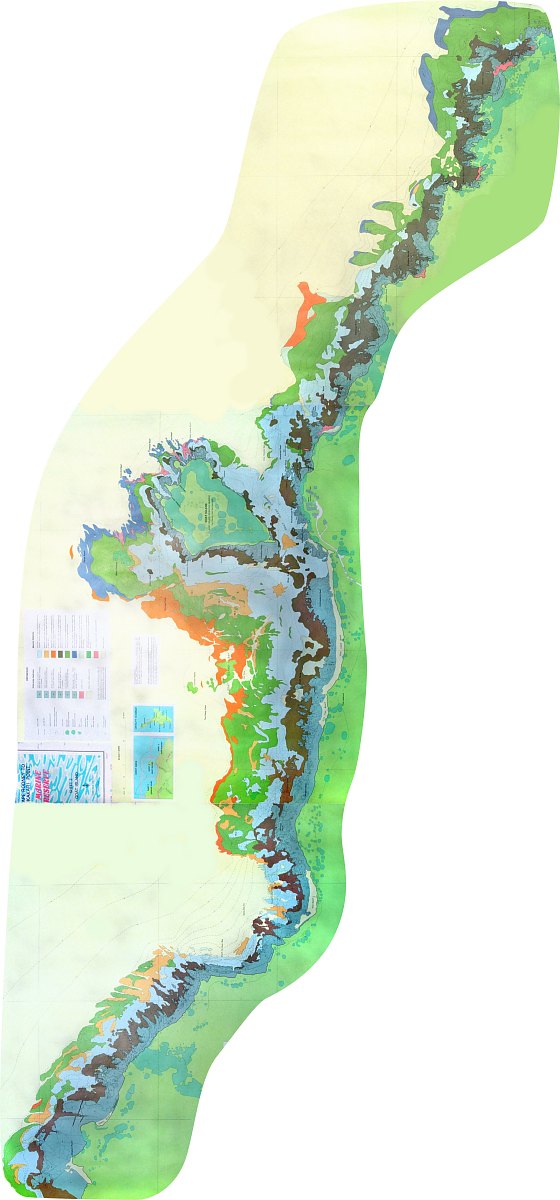
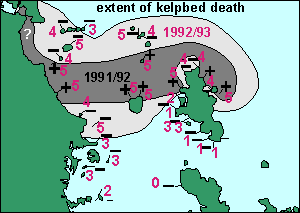 In
our own study we excluded the areas north of Whangarei after a preliminary
survey in March 1993. It allowed us to concentrate more on the areas surrounding
the outer Hauraki Gulf. Because of our extensive spatial data, we can say
that the bloom and its mortality did not centre on the Goat Island marine
reserve. See our map on right. The area around Port Fitzroy on Gt Barrier
I is rather patchy, possibly because this harbour has its own plankton
ecosystem and may have been spared some of the bloom's ravages. Also unaffected
water flows north and out of the Hauraki Gulf past these areas.
In
our own study we excluded the areas north of Whangarei after a preliminary
survey in March 1993. It allowed us to concentrate more on the areas surrounding
the outer Hauraki Gulf. Because of our extensive spatial data, we can say
that the bloom and its mortality did not centre on the Goat Island marine
reserve. See our map on right. The area around Port Fitzroy on Gt Barrier
I is rather patchy, possibly because this harbour has its own plankton
ecosystem and may have been spared some of the bloom's ravages. Also unaffected
water flows north and out of the Hauraki Gulf past these areas.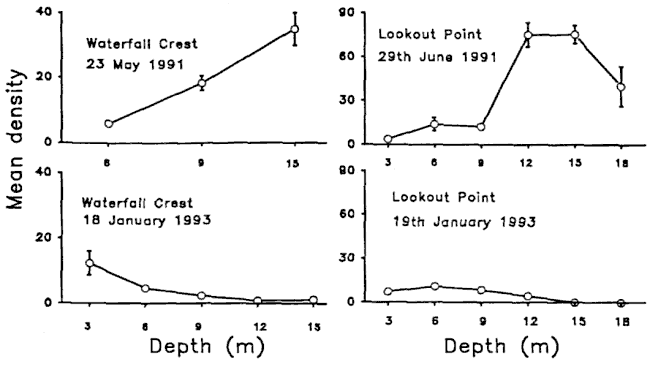
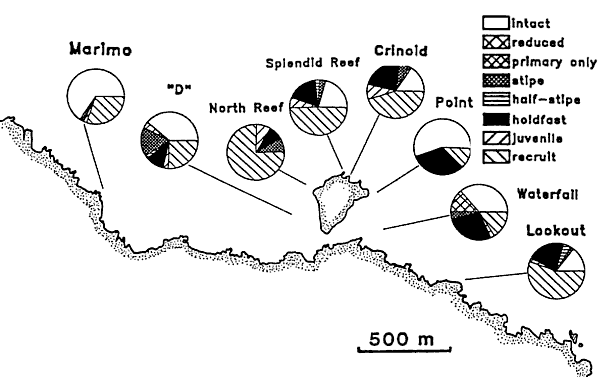
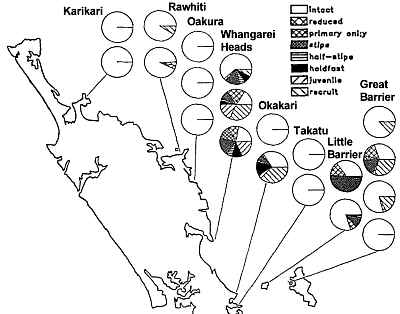 Regional
mortality patterns
Regional
mortality patterns
 The
1977 date is misleading because the situation stayed stable until the
large kelp dieback. Not a word about the large-scale kelp forest die-off
in 1993. The 1994 date is wrong since kelp recruitment failed that
year due to hungry grazers of all kind.
The conclusion is not at all
substantiated by the research done (see below). Large-scale disappearance
of urchin barrens was caused by the 1993 disaster. This aerial photo taken
in April 1999, shows the main barren area of the Goat Island marine reserve.
Notice how the kelp is beginning to invade, but most of the invasion happened
suddenly a year later. Scientists maintain that it was a gradual process
that began before 1980, which is proved false by this photo.
The
1977 date is misleading because the situation stayed stable until the
large kelp dieback. Not a word about the large-scale kelp forest die-off
in 1993. The 1994 date is wrong since kelp recruitment failed that
year due to hungry grazers of all kind.
The conclusion is not at all
substantiated by the research done (see below). Large-scale disappearance
of urchin barrens was caused by the 1993 disaster. This aerial photo taken
in April 1999, shows the main barren area of the Goat Island marine reserve.
Notice how the kelp is beginning to invade, but most of the invasion happened
suddenly a year later. Scientists maintain that it was a gradual process
that began before 1980, which is proved false by this photo.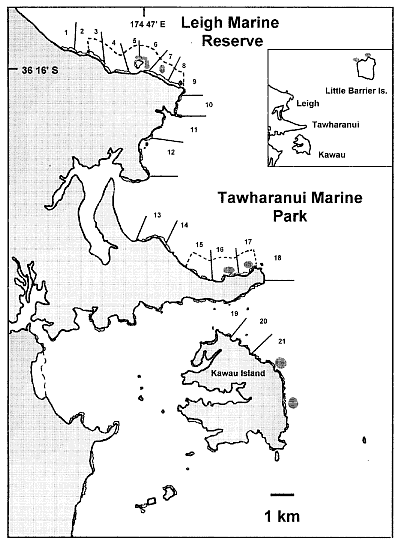 This
location map shows in dashed lines the countours of the Goat Island marine
reserve in the north and the Tawharanui marine park in the middle. The
numbered areas are the places where measurements were taken. The shaded
circles are the places where lobster abundance was measured.
This
location map shows in dashed lines the countours of the Goat Island marine
reserve in the north and the Tawharanui marine park in the middle. The
numbered areas are the places where measurements were taken. The shaded
circles are the places where lobster abundance was measured.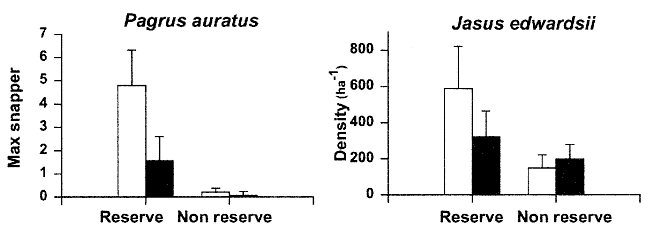
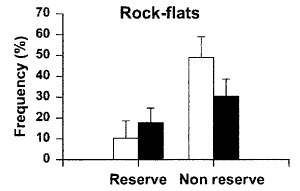 Algal abundance
Algal abundance
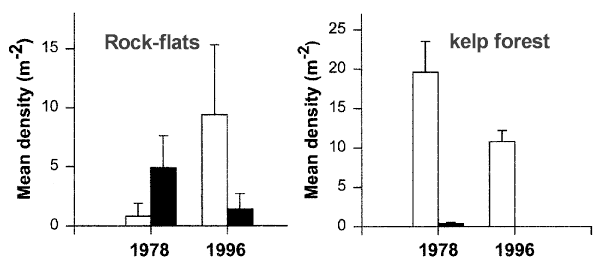
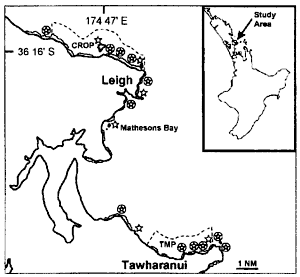
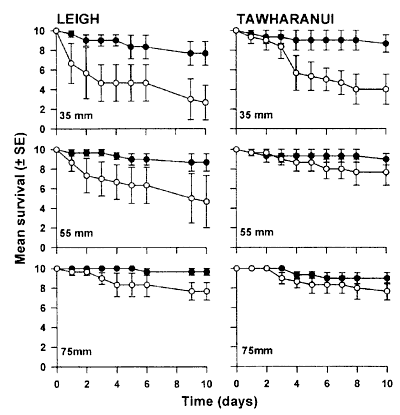
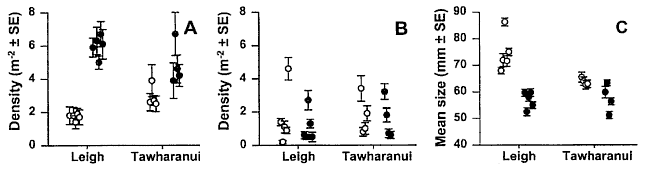
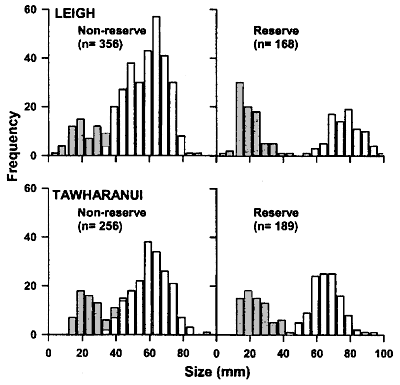
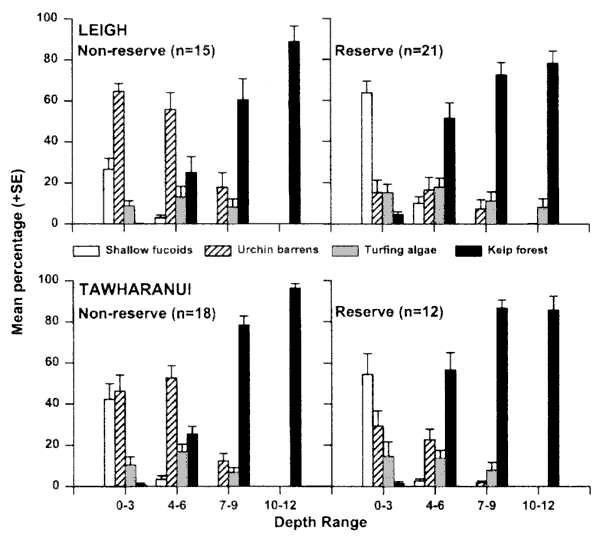
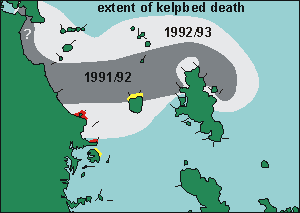 This
study does not give enough evidence for the above statement. When other
areas are taken into account (Kawau, Little Barrier, Great Barrier, Rakitu,
Mokohinau) it can be proved that this study is a failed experiment.The
map shows how the authors have observed kelp invasions inside two marine
reserves (red areas), but failed to observe similar effects elsewhere,
particularly in the yellow areas nearby the study area (yellow areas).
All areas where we observed kelp invasion of the urchin barrens were located
where previously kelpbed death has been observed. This can still be confirmed
today (2006).
This
study does not give enough evidence for the above statement. When other
areas are taken into account (Kawau, Little Barrier, Great Barrier, Rakitu,
Mokohinau) it can be proved that this study is a failed experiment.The
map shows how the authors have observed kelp invasions inside two marine
reserves (red areas), but failed to observe similar effects elsewhere,
particularly in the yellow areas nearby the study area (yellow areas).
All areas where we observed kelp invasion of the urchin barrens were located
where previously kelpbed death has been observed. This can still be confirmed
today (2006).