principles, physical and biotic factors and marine creatures
By Dr J Floor Anthoni (2007)
www.seafriends.org.nz/enviro/habitat/shore.htm
In the most inhospitable place of the sea,
where the hard sea shore is covered by the sea at high tide, and exposed
during low tide, lives a rich and varied community - why? The answer
to this question can be found only by understanding the living conditions
and limiting factors in this very narrow maritime zone. Meet the animals
and plants living here. Although the examples refer to the situation in
New Zealand, the principles behind them apply everywhere in the world.
If you don't live in New Zealand, you may still find these pages applicable
to your situation.
A vast number of principles to guide our investigations and observations.
Living in the sea and in the intertidal in particular, is spectacularly
interesting. (on this page, 14 pages)
The reasons why one species lives mainly here while another is found
there, does not only depend on physical factors but also on biotic factors
such as competition, predation and so on. However, the sea has some surprises
in store. (on this page)
Although the rocky intertidal is not a closed ecosystem, due to the
vast open sea bordering it, one still finds all methods of feeding here.
Also tips for doing a rocky shore study. (3 pages)
zoning The rocky shore is an ideal place to begin studying
the sea as it shows how different the sea is from the land. Many of these
differences are not obvious or not visible, and it pays to begin your study
with the introduction to marine habitats and our
chapter on biodiversity which
leads to a summary of differences between the marine and the terrestrial
world. On this page we'll encounter the principles applying to the intertidal
rocky shore, principles that apply everywhere in the world where a tide
forms an intertidal rocky shore.
When one traverses a snowy mountain in a mediterranean or tropical
climate from its top down to its base, one comes across several distinctive
zones, identified as follows:
permanent snow: the area with permanent snow has no plant life as
temperatures are too cold and the snow would lay over any plant or plant
seedlings.
snow-free in summer but snowed over in winter: as there exists no
sudden onset of summer or sudden ending, this is a zone with a substantial
gradient.
scree or broken rock: on the top of this zone one finds a minefield
of broken rock and sharp stones continually sliding down-hill. The rock
is broken here because of successive freezing and thawing which gives enormous
strength to water seeping into cracks and pores. This zone is not very
inhabitable because no suitable soil is found, but some lichens may survive.
alpine meadows: where the snow stays away for a suficiently long
time, and where soil develops, one finds a most diverse community of flowering
plants, mosses and lichens, in patches according to shelter, moisture and
warmth.
scrub forest: dwarfed and crippled trees dare to grow where the
temperature is insufficient for good growth, soils are frozen most times
and at times heavy snowfall is experienced. Wind speeds at this altitude
are also high. The scruffy trees grow to shoulder height but one can distinguish
some species that belong to the mountain forest.
montane forest: the mountain forest needs sufficient temperature
to grow a productive soil that may freeze over, as in a temperate winter.
Trees and shrubs may lose their leaves in winter as in winter-deciduous
(leaf-dropping) forests.
(sub) tropical forest: the temperature is always high enough for
good soil development and there is no need for dropping leaves in winter,
but in case the summer brings predictable droughts, some trees and shrubs
may be summer-deciduous.
coastal forest: the make-up of the forest is influenced by the sea,
such as by salt spray and strong winds.
In the above zoning example, obviously the snow/frost and the temperature
play an important role, and to some degree winds too. Also the amount
of sunlight may play a role. The main driving force is the yearly
season, and temperatures decreasing with altitude. So what would
the main factors be in the sea?
The
sea may experience the seasons too, being iced over at times in cool climates.
Ice over water reflects and absorbs most sunlight, thus seaweeds will die.
But most of the world's seas and coasts do not freeze over, although frost
may threaten creatures at low tide. Temperature and the amount
of light play a role for ocean productivity, but there does not exist
something like soil, except in very sheltered inlets (sand and mud flats).
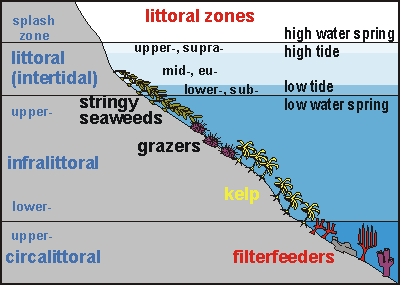
The diagram shown here shows typical coastal zoning in a temperate
climate. The intertidal (the subject of this chapter) is driven by the
tide. No tide, no intertidal, but there may exist a splash zone in case
of waves. Thus the intertidal zone is clearly defined by the movement of
the tide. Below it extends the infralittoral (photic zone) and circalittoral
zone (demarcated by lack of light to grow plants, or inadequate quantity
of light). These two zones are also clearly defined as they depend on the
clarity of the water. In murky water the photic zone may be as shallow
as 3m, whereas in a clear blue sea of 30m visibility, the photic zone may
extend down to 35m until also the quality of the light becomes insufficient.
But there is another zone inside the photic zone (infralittoral), demarcated
by the worst storms. Severe storms generate deep waves (swell) that can
cause severe damage even when the centre of such a storm passes 1000km
away from the shore. This powerful swell of some 200m deep (half of the
wave length of 400m), arriving at the continental shelf of 200m deep, experiences
the shallowing sea bottom as a hindrance, which absorbs the swell's energy.
As such deep waves travel further over an ever shallowing sea bottom, they
lose more and more energy. Thus the maximum energy that the most severe
waves can deliver to the coast, depends on the depth of the sea bottom.
The deeper the bottom, the worse the worst waves are. [note
that this has not yet been confirmed by mainstream science]. For
more about waves, read oceanography/waves/ In New Zealand, as in many other places, such devastating deep waves
cause a barren zone between the strong and flexible stringy seaweeds
and the more fragile stalked kelp. This zone is then occupied by
powerful grazers such as sea urchins (Evechinus chloroticus), abalone
or paua (Haliotis spp.) and Cooks turban shell (Cookia sulcata).
Such grazers are then able to prevent seaweeds from establishing themselves
in the barren zone, in effect keeping this zone barren, while also extending
it here and there. Terrestrial grazers such as sheep and cattle do something
similar on pasture and hoofed grazers (ungulates) in the tropical
savanna habitat.
The shape of the shore also depends on wave action, and like landscapes,
seascapes adjust their shapes for minimal loss in either erosion or opportunity
for life. Read our Least Loss
Landscapes hypothesis [which is not yet backed by mainstream
science]. The effect of this is that shores with massive wave exposure
(and thus deep sand bottoms) develop into vertical walls that bounce waves
back while absorbing only the least of their energy. Ironically, these
steep rock walls provide shelter as waves roll up and down
without much
shearing (currents). At the most sheltered shores we
find a shape that shears the wave at its top, as in a swimming pool's wave-absorbing
channel along all sides. Waves then shear over shallow platforms that are
covered in life, thus minimising erosion of the rock and damage to life.
It must also be noticed that the sunlight has a profound influence
on the shape of the shore, such that shaded shores (less life) are always
steeper under water than sunlit ones (abundant life).
The intertidal is defined by zones caused by the moon tide and sun tide.
When they work together, the tide is larger, which is called a spring
tide. In between occur the neap tides. For more about tides,
read oceanography/tides/. Note that
most places in the world have two tides each day, but there exist places
with only one tide each day, and even ones with no tide at all. Most places
have between 1 and 3m of tide swing, but places exist with up to 10m between
high and low tide.
Thus the intertidal zones can be defined clearly:
splash zone or maritime zone: the zone above high spring tide, that
is regularly sprayed or washed by waves. On exposed shores the splash zone
is wide, whereas on sheltered shores it becomes essentially absent. Above
the splash zone begins the terrestrial world, marked by lichens (white,
black, orange crusts) that are resistant to salt spray. Characteristic
of the splash zone is that sometimes for many days in succession, during
calm weather, the sea does not get there.
upper tide zone (supra littoral fringe): the water comes here only
twice each month during spring tides.
mid tide zone (eulittoral zone): the water comes here twice daily
but also retreats twice daily.
lower tide zone (sublittoral fringe): the water recedes from here
only twice each month during spring tides.
The diagram shows how the three zones are substantially altered by wave
action, which also blurs their distinction.
Compared to mountain zoning which covers several kilometres of altitude,
the zoning on the shore happens within a few metres. Also the living conditions
can change profoundly over short distances, due to irregularities like
ledges, cracks, stones and so on. Of course, waves can make this fine distinction
of zoning somewhat vague, and so can the shape of the shore. Here are some
new factors to consider:
topology: the shape of the shore determines very much its character.
A vertical rock wall for instance is not attractive for snails because,
once dislodged and falling into the deep, they cannot make it back. A shore
with rock flats is rich in grazing snails and other organisms, particularly
when it also has loose stones on it, but it may not extend to the upper
tide zone. Deep cracks and pools invite for high biodiversity.
rock hardness: the hardness of the rocky shore also plays an important
role. When the rock is soft , such as some sandstones, it erodes too quickly
for plants and barnacles to settle on, and even when they do, they are
all too easily dislodged. When a rock is very hard such as granite, many
organisms cannot attach because dissolution of the rock is part of their
method. A hard rock cannot be bored into by rock-boring clams such as piddocks
and date shells. Thus it remains unpitted and less attractive for other
organisms. Medium hardness rock which can be dissolved by acids appears
to invite highest biodiversity.
life protects: where the shore is hard enough for a permanent cover
of life, erosion suddenly becomes minimal as wave damage occurs to living
organisms that can repair the damage. Where degradation is rife to such
extent that it reduces the live cover, shore erosion suddenly accelerates.
Muddy water for instance, takes the light away for a protective (plant)
cover to grow.
wave-wash: waves make the intertidal a hell to live on. Where the
shore is deep and wave action during storms very strong, only few organisms
can survive (barnacles and stringy seaweeds). Most biodiversity is found
at medium wave action. In calm places one would expect even more biodiversity,
but pollution from the land in the form of mud, now makes life difficult.
Waves also blur the distinction between upper, mid and lower tide zones.
However, waves also have a positive rinsing effect, removing dust and sedimentation
and poisonous slime.
sand: some sand inside rockpools makes them more attractive for
sand-burrowing organisms like scavenging whelks, worms and some clams but
large amounts of sand can cause 'sand-blasting' in storms with loss of
life. Also large amounts of sand nearby can occasionally cover the shore
entirely, thereby killing almost all life.
fresh water: every rain during low tide showers organisms in fresh
water, which by osmosis tends to make organisms swell and die. For this
reason many organisms can seal their shells securely to the substrate,
avoiding this threat. Fresh water fills rock pools only at the very top
because fresh water is lighter than salt water, and is thus unable to cause
real hardship to those living beneath. Crustaceans and seaweeds are sensitive
to fresh water. Where fresh water intrudes, marine life recedes. Freshwater
springs are particularly life threatening in coral lagoons.
degradation: land-based degradation has caused seas to be 'sick'
from sediment and decomposing bacteria. In general, coasts near big rivers
suffer from degradation more than coasts with small water catchments. Seas
are 'sicker' near the main land and become 'healthier' further out towards
the edge of the continental shelf. Water quality affects shore biodiversity,
even though many animals are quite resilient. For more information, read
our decay section.
disasters: disasters occur occasionally. They can be natural such
as large storms or hurricanes or can be man-made such as oil slicks. Recovery
from a disaster is faster in a healthy sea than in a sick sea, reason why
even natural disasters have a man-made component.
One would have expected, that because all life begins with plants, most
of the intertidal would be covered in seaweeds, but it takes only one glance
to see that this is not true. Why?
It is caused by the massive influence of the sea outside, and the planktonic
food it contains (plant and animal plankton and their dead bodies). Thus
all organisms living from this source, can colonise the intertidal in competition
with seaweeds, a match that they win in their respective zones. This makes
barnacles, mussels and oysters invasive chalky colonisers as their abundant
plant source resides in the open sea. Within their chalky houses, these
animals are exceptionally equipped to prevent dehydration when the tide
is out.
surface meets bottom Planktonic organisms keep
themselves suspended either by swimming or by some buoyancy control mechanism.
But when the organisms die, they either sink down or float up. Thus the
two places where the food collects, are the surface and the bottom. It
so happens that the intertidal is the ONLY place where this happens, thus
planktonic food (phytoplankton, zooplankton and detritus) is aplenty. In
tidal inlets where wave action is weak, detritus abounds, whereas on the
exposed rocky shore, the surface 'scum' is a rich source of food.
But the surface can also
become a menace in the case of oil spills and other forms of pollution
that are surface-bound. This may explain the often sudden disappearance
of many organisms such as the red beadlet anemone
(Isactinia tenebrosa).
veges & pizzas When studying the organisms
of the rocky shore, one must distinguish clearly those that live from the
food produced on-site and those that live from imported food. The first
are like families living from their garden plots of vegetables; the second
are like families ordering their pizzas and fried chickens from far away.
The first live on low quality food (veges), the latter on high quality
food (proteins). The first cannot be as productive as the latter. Remember
though that even those living from locally grown vegetables, still benefit
from imported nutrients.
one cannot reproduce the rocky shore inside
an aquarium because the dominant influence of the open sea
cannot be simulated
One would also expect that intertidal zoning is fixed and predictable,
as the zonation diagrams shown above, suggest. But after some investigation,
one finds that this is not so. Why?
Of all the abiotic (physical) environmental factors discussed above,
the tidal range, exposure, shore shape and substrate are the most important
ones. Because each has its own 'sorting sequence', the resulting zones
are quite variable.
0701180: located behind a barrier rock, which shelters this
coast, stringy seaweeds are threatened by drying out at spring low tide.
The greenish zone is the mid intertidal. Pied shags are drying themselves
after a day's fishing. The reef flat is rich in organisms and tide pools
abound with seaweed species. The seaweeds are cartilage weed (Xiphophora
chondrophylla), flapjack (Carpophyllum maschalocarpa) and just
visible stalked kelp (Ecklonia radiata).
0701183: Never Fail Rock is an exposed rock in a 50m deep
sea). The spring tide covers no more than 2m (compare with the birds).
It is a very calm sea now at spring low tide and stringy seaweeds (Carpophyllum
maschalocarpa) are having a hard time. This is the sheltered side of
the rocks, and white bird excrement shows a curved splash zone where it
is washed away. The greenish zone is the eulitteral.
0701157: the intertidal in a healthy sea: high biodiversity
of seaweeds and other life. Every bit of rock is covered in long-lived
sea life. Stalked kelp dares to grow in the lower littoral fringe. East
coast Mercury Islands.
f218219: the intertidal in a degraded sea: low biodiversity.
Reef stars (Stichaster australis) surviving desiccation (drying
out) while reaching for the black band of flea mussels. Much of the shore
is not covered in long-lived sea life. No coralline pink paint! West Coast.
f009525: rock pools with seaweeds while all exposed rock
is covered in life, a healthy environment at Goat Island marine reserve.
This reef flat is found at the mid to lower littoral and there is no upper
littoral or splash zone. There are many broken rocks, some large, accommodating
high biodiversity. Featherweed and flapjack in the pools. Abundant coralline
pink paint.
f991016: a degraded rocky shore at low tide, showing little
variety and poor cover. The pink paint (Lithothamnion sp.) is almost
entirely absent, making the shore subject to rapid erosion. Long Bay marine
reserve. The white crusts consist of rock oysters and plicated barnacles.
f219610: a potentially interesting rocky shore with rocky
undulations capturing large rockpools and boulders littered everywhere.
Yet this is a very degraded shore with poor diversity and cover, due to
the release of treated sewage at the point in the distance. Whangaparoa
Peninsula. Very little pink paint.
f219722: a low tide platform at a very exposed shore has
gutters full of seagrass, caused by a high input of mud and nutrients.
Near Napier.
f222332: a seaweed washup after a storm, although natural,
can cause tremendous loss of life as seaweeds rot away and decomposing
bacteria kill sensitive life. A natural disaster such as this has a man-made
component: sea urchins died by eating a highly poisonous dinoflagellate
slime and this invited the kelp to grow in the urchin barren zone where
storms would take them out. The dinoflagellate Ostreopsis slime
was caused by unnaturally high nutrient loads in the water which is man-made.
f219317: intertidal zoning on a sheltered evenly sloping
shore continues in the vegetation above. Note how low the barnacle zone
is relative to the sublittoral fringe of brown seaweeds. Note also the
grey, orange and black lichens so close to the top of the intertidal zone.
Entrance to Whangarei Harbour.
principles Apart from the obvious physical factors that cause the zoning on the
shore, there are a number of other principles to consider, which makes
the study of the rocky shore so interesting.
roots/leaves: land plants need roots to find water in the soil and
to transport water and nutrients through 'pipes' (vessels) to their leaves.
They pump water up as water evaporates from their leaves. Land plants do
not like rock as they need soil for their moisture, nutrients and recycling
of decaying matter. In the sea it is an upside down world. Seaweeds do
not like sand because it is constantly on the move and seedlings would
not be able to survive. Seaweeds find their moisture and nutrients all
around, so they do not need roots or pipes and pumps for that matter. They
absorb nutrients while exchanging gases through their leaves. Seaweeds
can exist only on a substrate such as rock and their roots (holdfasts)
serve only to hold on to the rock. Note that eelgrass and mangroves (both
vascular
plants) do live in mud/sand, but only in very sheltered harbours where
their wandering roots (stolons) secure them against being washed
out. Note that botannically speaking, vascular plants have
leaves
but seaweeds, mosses, ferns and palms have fronds. But in this section
we will use the words leaf and stem freely.
waves: because water is 800 times denser than air, the force of
waves can be very destructive, but there are several survival strategies.
flexibility: For seaweeds it doesn't pay to be rigid like
a tree but to yield to the water movement instead. The stringy bladderweeds
follow this principle. And just in case there are no waves, they have gas-filled
float bladders to keep them upright. Note that the stalked kelp is kept
upright by its stiffness, even though its stalk can flex and yield to water
movement. Observe how all bladderweeds with flat stems have a corkscrew
'foot' that allows them to bend in all directions. Note that trees have
both rigidity (in stems and branches) and flexibility (in leaves).
streamline: where the water rushes fast over the rock (shearing
force), it pays to have a streamlined shape. Limpets and chitons have such
a shape as they are flat, with a large suction foot. But such a shape cannot
move and turn easily. It also claims a large patch of the shore, just for
sleeping. This shape is also worst for being stepped upon. When humans
walk on the shore, their weight pushes down onto the shore through a small
surface under the shoe. Limpets never evolved with such a threat, reason
why people can easily break their shells. This is one reason it is desirable
to visit the rocky shore on bare foot.
strong shell: a strong shell protects not only against being sand-blasted
but also against being rolled around when dislodged. A strong shell also
protects against predation as it can less easily be broken or drilled into.
Many shore creatures have strong shells, particularly those living on exposed
surfaces.
large foot: particularly the grazing snails have a large foot to
secure themselves.
small size: being small reduces the water's drag, but more importantly,
enables creatures to find a sheltering crack or pit.
water breathing: in order to survive being submerged for hours,
organisms need to be able to breathe water through gills. Although some
mammals (dolphins, whales) and other terrestrial animals (turtles, snakes)
can be amazing in their breath-holding, they eventually need to surface
and none would be able to last attached to the intertidal.
uni-directional flow: it is interesting to note that water breathers
arrange for a flow of water over their gills in one direction. They cannot
afford the extra energy to stop the water, and accelerate it in the reverse
direction, such as all air breathers do. Air is 800 times lighter than
water and its flow can be reversed without much loss in energy.
cold-blooded: an animal that breathes water cannot be warm-blooded
because the water comes in close contact with the blood, over a large area
(gills). There would not be enough energy to stay warm. All submerged animals
must therefore be cold blooded (poikilotherm) and this allows them
to shut down over considerable periods.
on stand-by: cold blooded animals are able to go on stand-by (shutting
down) because they do not need to expend energy staying warm. For instance,
the difference between a human sleeping (80-100Watt) and working hard (300W)
is not large. When 'shut down' for the night, humans still expend a considerable
amount of energy on warmth. The ability of going on 'stand-by' allows organisms
to stay out of water for considerable time periods, some even up to two
weeks (barnacles, periwinkles). The moment they become inundated, they
are then able to switch on and continue their meal.
drying out: drying out (desiccation) is one of the severest
threats to those staying out of the water. To counter this problem, organisms
have developed several strategies:
survival pack: organisms store extra water. Snails lock a few drops
of water up over their gills and insides. Through this water they can still
breathe a little. By quickly removing a snail from the rock and turning
it over, one can see it withdrawing deeper into its shell while sacrificing
its 'survival pack'. Seaweeds store extra water by being puffed up (crusting
seaweeds and others). The survival pack also helps against overheating
by allowing some of the water to evaporate, which causes cooling.
huddling: by huddling together, organisms can retain more moisture
than each in isolation. They also shelter one another against drying winds
while also minimising exposure to sunlight.
finding cracks: cracks in the rock and any discontinuity (sudden
change) in the surface, invites hairy algae to grow larger than usual because
grazing them is difficult. These algae lock up moisture, which helps other
organisms to survive. Cracks can also provide shade against direct sunlight.
finding shade: avoiding direct sun exposure by seeking the shaded
side of a rock helps considerably.
hiding under stones: stones offer the perfect shelter from wind
and sunlight while also providing a solid roof against browsing seabirds.
That is why turning stones when studying the rocky shore, provides the
most unusual finds. The larger the stone, the more likely it will stay
in place amidst wave action, and the more diversity can be found underneath,
and also older organisms.
heating/cooling: once exposed to the air, intertidal organisms are
affected by the sun's heat and the drying effect of wind. On a winter day,
organisms can become frozen before the tide returns. Survival strategies
against overheating and cooling are similar to those against drying out,
see above. To counter extreme frost, some marine creatures have 'anti-frost'
compounds in their blood.
increased metabolism: many sea creatures are seen 'basking' in the
sun in shallow water (sharks, pelagic fish, sting rays, and many more).
This is not for tanning or for feeling the heat of the sun but to warm
up. As the surface layer can be 4-6ºC higher in summer, it improves
metabolism (food digestion & growth) and could also bring a sense of
well-being. Thus creatures who 'bask', grow faster and make more spawn
than those who don't. Likewise, being able to stay out of the water, and
thus able to warm up, could mean a higher growth rate and egg production.
oxygen release: gases like oxygen and carbondioxide dissolve well
in water, and more so in cooler water. Thus when water warms up, gases
tend to escape, which means that their availability increases. Fish seek
warming waters for their increased oxygen levels, which may also increase
their metabolism.
exposure to degradation: we discovered that degradation in the sea
is caused mainly by planktonic decomposing bacteria that are capable of
causing infection. The chance of infections is proportional to the time
being exposed to it. Thus being able to live outside the water for some
time, reduces the chance of dying. It is a good idea to spend a short time
submerged to feed, followed by a long time to digest the food. For more
on degradation, visit the decay section.
salinity: when sea water evaporates, it becomes more salty, which
accelerates fluid loss from organisms through osmosis (water moving
from a weaker concentration in the body, to a denser one outside). Rockpools
may become too salty to survive in. But all marine organisms need to 'drink'
salt water while excreting salt, in order to stay moist, so added salinity
is easy to cope with for animals, although less so for plants.
size: size matters considerably for survival. Being small reduces
the drag of the waves, requires less energy to maintain, while one's weight
is easier to carry in the presence of gravity (out of the water). Rockpool
fishes are from necessity small. Being small is a disadvantage against
predation as escape is too slow and a thick shell unaffordable. But predators
who have to be larger than their prey, are disadvantaged by being less
capable of surviving out of the water or on wave-exposed surfaces.
weight/gravity: while submerged, most organisms are almost weightless.
But being out of the water requires them to carry their full weight, reason
why they do not like to move around. An octopus for instance, a strong
mollusc, is hardly capable of movement above water. Many molluscs like
grazing snails, limpets, chitons and abalone are remarkably strong because
they need to resist strong water movement. But being attached strongly
goes against movement. Thus they either move while lightly attached, or
stay put while strongly attached.
hydraulics: many underwater creatures (molluscs, echinoderms) master
a mysterious force, that of hydraulics. A snail withdraws into its shell
by expelling water and shrinking. For instance, it extends a siphon to
half its body length by hydraulic forces. It sucks to the rock like a suction
cup, and once 'stuck', requires very little energy to maintain suction.
This enables them to remain stuck firmly in place while 'on stand-by'.
A snail or clam can burrow straight into the sand by mysterious techniques,
of which hydraulics is a part. It can also suddenly emerge from under stones
while lifting these with ease. It can disappear in a crack or under a stone
by shifting its shape to suit. A weight-less army tank would not be able
to do any of this.
oxygen: in order to survive, animals need oxygen and plants carbondioxide.
But both can go on 'stand-by', surviving long periods of oxygen starvation.
Note also that above water there is still oxygen and CO2, although less
easily available.
feeding: for most intertidal animals, feeding stops once the tide
goes out.
fishes: rockpool fishes claim their large territories when the tide
is in, and they retreat to rock pools which they share with their rivals
in a kind of 'truce'. When the tide is in, they show how remarkably adapted
they are to wave-wash, with their large breast fins clinging onto the rocks
and speeding to and fro in the shallowest of water.
grazing snails: when the tide is out, they stop feeding and go on
stand-by. Only inside rockpools can one find grazing snails, limpets and
chitons.
echinoderms: some starfish are able to survive out of water (cushion
star). They stay moist by keeping water between their frilly tubefeet on
top. On a sand flat they are able to transport water by means of their
tubefeet, from underneath to their backs. Submerged echinoderms like sea
urchins and brittle stars continue feeding as normal.
predation: also predation stops when the tide is out, but once on
its prey, a predator will not let go, because the meal may take a week.
Predators such as the dark and white rock shells (Haustrum sp.)
have a large space under their shells which acts as a mitten (glove) to
hold the prey and a survival pack of water, while allowing them to attach
as well.
body plans: more so than the terrestrial environment, the sea has
a much larger 'arsenal' of body plans in the form of ancient designs that
still occupy unique and often important functions today. The body plans
are reflected in the various phyla, the main groups of life, some
of the most obvious in the animal kingdom are mentioned here:
Porifera: the sponges are sessile animals that protect themselves
by being inedible or rightout poisonous or by having spiky glass skeletons.
Sponges in general grow only slowly and need to be submerged.
Cnidaria (=Coelenterata): the flower animals with stinging cells.
These are the coral builders in the tropics but in the temperate climates
play a lesser role: anemones, gorgonians, solitary corals, jellyfish. The
flower animals have a very simple form: just a stomach with a mouth, surrounded
by tentacles. They are very soft and like to stay under water.
Ectoprocta (=Polyzoa=Bryozoa): the mat forming moss animals are
tiny flower animals living inside tiny boxes, complete with lid. These
boxes join up in regular patterns. They too like to remain submerged as
under stones.
Brachiopoda: the ancient lamp shells that look like molluscs with
two shells.
Annelida: the round worms and bristleworms that play an important
role in decomposing dead animals. They also include the plankton-feeding
tube worms. Worms form an important food source for others.
Arthropoda: the joint-legged ones, of which the Crustaceans
are the most common on the shore: shrimps, crabs. Also the numerous but
largely invisible sea slaters and sand hoppers. Crustaceans live
inside an external skeleton and in order to grow, must discard their old
skeleton to grow a new, bigger one. This is called moulting. Having many
legs leads to diversification, reason why crustaceans have adapted to many
kinds of feeding.
Mollusca: the soft-bodied ones including inkfish, shells, clams,
chitons, limpets and nudibranchs. Molluscs do not have fixed shapes and
can adapt to the size of the environment. They represent a huge diversity
in the sea. Most molluscs are hermaphrodites, being male and female
at the same time. This enables them to live in seclusion while multiplying
without mates.
Echinodermata: the prickly skinned ones: sea urchins, starfish,
sea pens and sea cucumbers. Echinoderms also live inside an external skeleton
made of many individual plates, separated by living tissue. They can grow
by growing each plate separately. They can also shrink in size to suit
the amount of food they can get. Some sea stars can reproduce by splitting.
Having many tubefeet, they too have adapted to a wide range of food: grazing,
scavenging, predating, detritus feeding, plankton feeding.
Chordata: the ones with a spinal cord. Of course mammals, birds,
reptiles and amphibians belong to this phylum, but also the Classes:
Ascidiacea: the ascidians or sea squirts. These live from phytoplankton
by sieving the water through a fine bag.
Osteichthyes: the bony fishes: the little fishes found on the rocky
shore. There is no space for big ones.
species interaction Whenever a species finds a place favourable for living and reproducing,
it discovers that other species think so too. Competition begins. After
a long time of evolution, species differentiate such that each survives
on its particular skills that the other does not have. Thus evolution tends
to minimise competition. Likewise, predators develop skills to catch and
eat others, each predator species with slightly different skills so that
they do not need to compete. At the same time the prey develops better
defences.
The science of ecology teaches the following classical species interactions:
neutral: not benefiting or damaging either species
competition: competition for space, for food, for
shelter.
association: one species is usually but not necessarily and not
exclusively associated with another. For instance, barnacles make the surface
rather rough and unsuitable for medium-sized grazing snails, but little
ones can negotiate the rough terrain and even find shelter inside empty
barnacle shells, even when they are not normally found at this altitude
without barnacles.
negative: benefiting one but damaging another
predation: predators eat others, thereby killing an individual and
reducing the numbers in a population or community. Yet predation can be
beneficial because often the weak and sick are targeted, and they help
limit populations before these run out of food, which can cause unpleasant
death while the health of the population weakens.
parasitism: a parasite benefits from exploiting its host, whereas
the host loses.
positive: one species benefits another and vice versa
commensalism: occurs when an individual obtains a benefit from a
different species without damaging it.
mutualism: occurs when an individual obtains a benefit from another
species and, at the same time, the second species obtains a benefit from
the first one. Mutualism is not obligated, which makes it different from
symbiosis.
symbiosis: species benefiting one another but also depending on
one another. If one of the symbiotic individuals perishes, the other
also perishes by losing the source from which it was obtaining a benefit.
Interactions within a species population (intra-specific interactions)
can be:
social dominance: the stratification of groups into a society given
by the influence that one individual or one group of individuals has on
the other individuals or groups into the same society.
social hierarchy: the stratification of the individuals that consists
in the domination that an individual has on the other individuals of the
same population.
territoriality: the demarcation and defense of a physical area that
is defined by an individual or by a group of individuals.
intraspecific competition: happens when two or more individuals
of a population try to obtain a factor needed by all individuals from the
environment where they inhabit.
effectively: if the competition is brought to a harmful struggle
between two of the stressed individuals of a population
unintentionally: if the competition does not imply a deadly or harmful
ritual, but a natural application of abilities to achieve a required factor.
agonistic behavior: if the contest implies a risk-free ritual of
threatening and submissive behavior
ritualistic competition: if the competition does not imply a ritual
Obviously, most of the inter-species interactions mentioned above relate
to higher organisms, except for competition where the amount of food and
the distance to neighbouring competitors is critical.
One
method of scientific study of the intertidal zone is by laying out a measuring
tape and measuring species densities along the tape. The species counts
can then be depicted in the form of a kite diagram as shown in this
example of grazing molluscs. The width of the black kite represents the
number of individuals found within a defined quadrat. (Note, however, that
the kite diagram shown here is more conceptual than representing actual
numbers.) Students are then asked to explain why certain species are found
where they are, why some overlap with one another and others don't, using
all the knowledge explained before. In our example, all species shown are
grazers which suggests that competition should be a major biotic factor
as well as exposure to desiccation (drying out).
However, also some important information is missing. To begin with,
it does not show where the high and low tide marks are. One should also
know that this is a shaded and reasonably sheltered shore, reason why the
grazing slug Onchidella is found here, crawling out of their hideouts,
grazing the sea weeds above. So the zone builders not shown on the diagram
are important biotic factors as well. For instance Siphonaria is
usually associated with the blood crust Ralfsia or Gelidium.
One should also know that this is an almost vertical shore, reason why
Cellana
ornata is found and Melagraphia is lower down than usual. The
message here is that there are many hidden clues before one can understand
the ecology behind this diagram of grazers. But there is more to consider.
If one were to study the same area of shore over a long time period
(longer than 10 years), one may notice that the kite diagram changes seasonally
and annually, and often quite suddenly too. So in the end, our entire toolbox
of ecological factors proves to be of no use. None of the explanations
we can muster holds true over a sufficient span of time. Why? Why does
ecology in the sea appear to make no sense?
The most important property of the sea is
that it is strange; stranger than we can imagine; nothing works the way we anticipate.
There are a number of invisible ecological factors that we do
not find on land:
imported food: we mentioned this before. Much if not most of the
food input to the rocky shore comes from places far away as an inexhaustible
supply. For barnacles who feast on dead and living zooplankton therefore
food is not limiting; only space is. Even grazers like limpets take in
a lot of nutritious detritus supplied from afar. Slime tracks from snails
can serve to trap detritus, eaten by same snails, or which bacteria can
convert to nutrients and so on.
imported nutrients: for algae living on the shore, there is a never
limiting supply of new nutrients. For them mainly the amount of sunlight
and warmth are limiting.
bacteria as food: only in the soil can one find organisms living
from bacteria, but in the sea the bacteria can assume the plankton's main
biomass. Furthermore every surface (even fish skins and seaweed skins)
is covered in dense mats of bacteria that can be eaten. Nearly all surface
grazers consume the fast growing bacteria as substantial parts of their
diets.
bacteria as killers: one of the largest ecological factors in the
sea, and so far entirely overlooked by mainstream science, is the planktonic
decomposer guild (an association of heterotrophic bacteria) that also brings
disease. Sea water is not only thin nutritious soup but also thin infectious
sewage. Living in the sea is a delicate
balancing act, and living in the intertidal does not exempt. Bacteria
can suddenly change the goal posts, causing major habitat shifts and mass
mortalities. [see our discoveries
that have not yet been confirmed by mainstream science, and principles
of degradation]
broadcast spawning: on land the offspring are born near their parents,
so we can understand how populations grow (gradually and predictably) and
describe that in mathematical formulas. Sea organisms however, produce
excessive amounts of spawn, most of which serves as food for others. So
the number of offspring born on suitable habitat is entirely unpredictable.
From one day to the next, a shore may become overwhelmed with the offspring
of a single species. Likewise, the offspring from a mollusc may be born
hundreds of kilometres away.
offspring from successful survivors are not born where their survival
skills matter: this is related to the previous factor, which makes
evolution and adaptation on land faster than in the sea. this leads to
the following:
all seas are connected whereas on land there are isolated pockets
like islands, and land separated by rivers, ice and mountain ranges.
visitors from other habitats: on land only birds who can fly long
distances can become visitors from other habitats, but the rocky shore,
even the estuarine flats, are so narrow that visitors who do not live there,
are common and can have a decisive influence. Wading birds on mudflats
are a good example, but with a rising tide come a large array of marine
visitors who have only one thing in mind: exploitation. Even little triplefins
(small fish without float bladder) become redoubtable predators, picking
off all newly settled barnacles while operating from a rockpool many metres
away. Visiting herbivorous fish (weed eaters) may in one visit have more
influence than all the resident grazers together all week. The intertidal
habitat is incredibly narrow.
failure is more important than success: in terrestrial (land-) ecology
we think in terms of survival of the fittest, which sprouts from
the foundation that offspring are few and born near their parents. From
this observed fact we then explain behaviour designed to propagate the
genes of the fittest (best suited). On land, survival is important.
In the sea with broadcast spawning, long planktonic larval stages and larvae
drifting on currents, this paradigm (way of thinking) is no longer defensible.
On
the intertidal shore where life is brutal and short, failure becomes the
dominant selector. In other words, on land the fittest are selected
by survival during life, resulting in advantage during reproduction, whereas
in the sea they are deselected by failure during birth (recruitment). So
we can understand the intertidal (and the sea) only from this perspective.
Look
at intertidal zoning and try to understand it in terms of failure as the
big selector! [Reader please note that this point is
highly counter-intuitive and controversial and is perhaps unsuitable for
teaching]
The result of all this is that the intertidal shores (and somewhat the
sea) are rather counter-intuitive (paradoxical), resulting in patchiness
(randomness) with large variations over time.
Now we've come full-circle with the most important ecological factor
saved for last. Read this chapter again, but now from the paradigm (way
of thinking) that failure (death) is more important than success (survival).
Also consider this:
The most overlooked ecological factor is
TIME; when we look at a situation for long enough,
our knowledge becomes a riddle. - Floor Anthoni 2009
Further reading References in blue are available
from the Seafriends Library
Adams, Nancy M (1994): Common seaweeds of New
Zealand. Canterbury University Press.
Adams, Nancy M (1994): Seaweeds of New Zealand.
Canterbury University Press.
Ayling, Tony & Geoffrey J Cox (1982): Collins
guide to the sea fishes of New Zealand. Collins.
Cometti, Ronald & John Morton (1985): Margins
of the sea: exploring New Zealand's coastline. Hodder&Stoughton.
Dell, R K (1963): Native crabs. (Nature
in New Zealand series). AH & AW Reed.
Enderby, Jenny and Tony (1998): Goat Island
marine reserve, Leigh, New Zealand. Enderby.
Fell, Barraclough H (1962): Native sea-stars
(Nature in New Zealand series). AH & AW Reed.
Forest J et al. (2000): Paguridea (Decapoda
Anomura) exclusive of the Lithodidae. NIWA Biodiversity Memoir 114
Foster, Brian A (1978): The marine fauna of
NZ: Barnacles (Cirripedia: Thoracica). NZ Oceanographic Inst Memoir
69.
Francis, Malcolm (1988-2001): Coastal fishes
of New Zealand - an identification guide (3rd edn). Reed Publishing.
Gunson, Dave (1984): Collins guide to the
New Zealand seashore. William Collins.
McLay, Colin L (1988): Crabs of New Zealand.
Leigh Laboratory Bulletin 22.
Morton, John and Michael Miller (1968): The
New Zealand sea shore. Collins.
Morton, John (2004): Seashore ecology of New
Zealand and the Pacific. David Bateman.
Paulin, Chris & Clive Roberts (1992): The
rockpool fishes of New Zealand. Te ika aaria o Aotearoa. Museum of
New Zealand.
Penniket, J R and Geoff Moon (1970): New Zealand
seashells in colour. AH and AW Reed.
Powell, A W B & B J Gill (Ed) (1947-1998):
Powell's
native animals of New Zealand (4th edition). David Bateman
Powell, A W B (1979): New Zealand mollusca;
marine land and freshwater shells. William Collins.
Schiel, David R (2006): Guide to common intertidal
species of the South Island, New Zealand. University of Canterbury.
Stace, Glenys (1998): What's around the rocks,
a simple guide to the rocky shore. Viking.
Walsby, John (1990): Nature watching at the
beach. Wilson & Horton.
What's new? yyyymmdd
20090414 - species
interaction added with hidden ecological factors
20070710 - Pages
made smaller for ease of access and printing.
20070703 - All links on this page were corrupted
but have now been corrected. Oops.
20070330 - Further
refinements and more photographs.
20070318 - Final
touches but many photos still to come slowly. Published on the Web.
20070302 - Seaweeds
part3
published, but more work needs to be done.
20070226 - First
published on the Web, including identification part1
and part2.
 The
sea may experience the seasons too, being iced over at times in cool climates.
Ice over water reflects and absorbs most sunlight, thus seaweeds will die.
But most of the world's seas and coasts do not freeze over, although frost
may threaten creatures at low tide. Temperature and the amount
of light play a role for ocean productivity, but there does not exist
something like soil, except in very sheltered inlets (sand and mud flats).
The
sea may experience the seasons too, being iced over at times in cool climates.
Ice over water reflects and absorbs most sunlight, thus seaweeds will die.
But most of the world's seas and coasts do not freeze over, although frost
may threaten creatures at low tide. Temperature and the amount
of light play a role for ocean productivity, but there does not exist
something like soil, except in very sheltered inlets (sand and mud flats).











 One
method of scientific study of the intertidal zone is by laying out a measuring
tape and measuring species densities along the tape. The species counts
can then be depicted in the form of a kite diagram as shown in this
example of grazing molluscs. The width of the black kite represents the
number of individuals found within a defined quadrat. (Note, however, that
the kite diagram shown here is more conceptual than representing actual
numbers.) Students are then asked to explain why certain species are found
where they are, why some overlap with one another and others don't, using
all the knowledge explained before. In our example, all species shown are
grazers which suggests that competition should be a major biotic factor
as well as exposure to desiccation (drying out).
One
method of scientific study of the intertidal zone is by laying out a measuring
tape and measuring species densities along the tape. The species counts
can then be depicted in the form of a kite diagram as shown in this
example of grazing molluscs. The width of the black kite represents the
number of individuals found within a defined quadrat. (Note, however, that
the kite diagram shown here is more conceptual than representing actual
numbers.) Students are then asked to explain why certain species are found
where they are, why some overlap with one another and others don't, using
all the knowledge explained before. In our example, all species shown are
grazers which suggests that competition should be a major biotic factor
as well as exposure to desiccation (drying out).