Myths and fallacies exposed,
part 7 Dissected by Dr J Floor Anthoni (2003)
www.seafriends.org.nz/issues/cons/myths7.htm
In this article the author attempts to come clean
by admitting that those in favour and those against marine reserves, can
have their facts wrong. He does so by presenting what is available as reliable
scientific knowledge in order to dispel the myths. However, in doing so,
he also falls into a number of traps as we will identify. For a scientist,
such carelessness is unforgivable because the public must be assured that
scientists pursue truth. For those interested in the falsities within marine
science and the marine reserves debate, this document forms an interesting
study. An important outcome is that three frequently told myths are now
officially biting the dust.
This article appeared in PDF format and was translated
unabridged into HTML for ease of access. In the process, the figures and
diagrams have suffered somewhat in quality. The original text appears in
black, and our comments in blue.
The New Zealand Marine Reserve
Experience: the science behind the politics Russ Babcock (2003) University of Auckland Leigh Marine Laboratory
PO Box 349 Warkworth, New Zealand
present address
CSIRO Marine Research Floreat, Private Bag No.5, Wembley WA 6913, Australia
russ.babcock@csiro.gov.au
ABSTRACT
Debate surrounding the effectiveness, or otherwise, of marine reserves
has not been well informed by data. However, in areas where marine reserves
have been established for some time, valuable information is now becoming
available. New Zealands no-take marine reserves have demonstrated large
increases in abundance and size of exploited species such as Snapper Pagrus
auratus, Spiny Lobster Jasus edwardsii and Blue Cod Parapercis
colias in marine reserves. Significant increases have been rapid, occurring
within one year in the case of snapper, but only evident where full no-take
protection is afforded. These increases in biomass of exploited species
translate into levels of egg production between 4.4 and 18 times those
of surrounding areas of coastline.
The recovery of a fishery within a closed
area is not disputed and indeed the fact that this happens so quickly,
proves that the damage done by fishing was little. Likewise, the benefit
of a marine reserve cannot be expected to be large.
There is no evidence of increased egg production translating into increased
recruitment to fished populations, but such effects would be impossible
to detect given the small proportions of coastline protected in reserves.
There is some evidence that, in marine reserves, benthic soft bottom communities
have responded to protection from direct effects of fishing such as trawling
and dredging.
New Zealand has over 3000km2 (twenty
times the area now in marine reserves) in cable ways and ammunition dumps
which have enjoyed de-facto protection as no-fishing areas, but scientists
never studied their benefit to fishing, recruitment and larval production.
The public sees this as a serious omission. Where is the evidence of the
claim that soft bottom communities inside reserves have 'responded'?
More surprising have been the indirect responses of benthic reef communities
to protection from fishing. Recovery of predators such as P.auratus
and J.edwardsii has allowed urchin-dominated barrens areas to revert
to more highly productive kelp forests. In this way reserves have allowed
us novel insights into ecosystem function as well as the pervasiveness
of indirect fishing effects.
The scientific work mentioned here is
the most shoddy research produced in the recent quarter century and has
been rebutted extensively as being no more than a myth, worse still, it
is the indirect effect of environmental degradation. Why the author keeps
hammering this shows how persistent a myth can be. On the Seafriends web
site visit Science Exposed and our own study,
Hauraki
Gulf Marine Survey 1993. The fact is that the extensive kelpbed death
of 1992/93 caused subsequent death of sea urchins and an invasion of their
barrens by seaweeds. The same happened also outside the marine reserve
in various locations, particularly at Little Barrier Island, some 20km
away.
New Zealand reserves offer no direct evidence of the often-touted spillover-related
enhancement of fisheries yield. However, they also show that reserves do
not lock up fisheries resources and at least for J.edwardsii,
CPUE (Catch Per Unit Effort), yield and costs are just the same adjacent
to a reserve area as in open fishing areas nearby. Thus both conservation
goals are achieved at no cost to the fishery. [oops!]
Spill-over from marine reserves is one
of the main myths propagated by marine scientists. Those fishing the sea
know it is not true because they have extensive knowledge of where the
fish roams. The above statement therefore is very important, coming from
a marine scientist and backed by scientific fact. However, the author is
seriously mistaken claiming that reserves do not lock up fisheries resources.
What about the lost fishery inside the reserve? When 20% of the
sea is locked up this way, a similar part of the total fishery is lost
for future generations. One does not need to be a rocket scientist to understand
this. In the case of NZ, this would amount to over $200 million per year!
How is it possible to claim the last sentence? Are scientists so naive?
Despite advances in fisheries management structures in New Zealand, such
as the Quota Management System, significant uncertainty remains about levels
of stock abundance and catch rates. This is true even for New Zealand s
best-studied stocks, such as P.auratus. Given that even the best
fisheries management systems remain demonstrably less than perfect, it
seems reasonable to try and guarantee some minimum level of stock abundance
by putting in place marine reserves. On balance there is ample evidence
to show that positive outcomes can be provided by reserves, and little
or no support for suggestions that reserves will have negative effects
for both conservation and fisheries.
It is known that fisheries management
is less than perfect, as knowledge attempts to catch up with discoveries
of new stocks. But the QMS is guided by sincere discussion and debate between
commercial and recreational fishermen (which cannot be said of marine reserves).
These are the people who stand to lose from fisheries collapses. By comparison,
the existing marine reserves in New Zealand have an even worse track record
of failure. At least two out of three of our coastal marine reserves have
failed because they are degrading from bad to worse. They do not protect
the marine environment. The fact that scientists have turned a blind eye
to the sea's worst enemy, land based pollution, shows how myopic scientists
can be and how irrelevant their research and statements in fact are, as
is also evident from this paper.
Opponents to marine reserves do not claim that
marine reserves are detrimental to the environment, but that their benefits
have been grossly exaggerated, and that there are better ways to save the
sea.
Introduction The idea of marine reserves, marine parks or marine protected areas
(areas of sea where some or all of the normal range of extractive activities
are prohibited) has existed in one form or another since the mid twentieth
century. However, the level of discussion surrounding this means of protecting
the marine environment has increased remarkably since about 1990. This
reflects increasing levels of general environmental awareness, the high
profile of such marine protected areas as the Great Barrier Reef World
Heritage Area, and the gradual recognition of the scope of human impacts
(particularly fishing) on marine ecosystems (Pauly et al. 1998). The public
debate among those who would like to retain the status quo (Lough 2002)
and those who believe in the need for the rapid and widespread implementation
of new approaches, such as marine reserves (Roberts and Hawkins 2000) has
taken place at many levels. At one end of this spectrum of views are scientific
institutions, such as American Association for the Advancement of Science
which has promoted frequent articles in journals, such as Science, discussing
the benefits of marine reserves (Schmidt 1997). At the other are community
meetings and local print and electronic media around the world where concerned
members of the public, conservation groups and a range of commercial and
recreational fishing interest groups attempt to promote what they perceive
as the merits or demerits of setting aside parts of the sea.
The world has over 1200 marine parks
and reserves, of which more than two out of three have failed. There exists
a clique of marine reserves scientists who push the idea of marine reserves
too far beyond what these can do well. In their largely opiniated articles,
not backed by scientific evidence, they exaggerate on the one hand the
threats from fishing and on the other hand the benefits from marine reserves.
They do not distinguish between bio-geographical areas (like what applies
to NZ and when) and they do not acknowledge well managed fisheries. The
above mentioned articles are typical. It is an indictment of science that
organisations such as AAAS do not exert sufficient criticism to separate
facts from myths and fallacies. Read for instance their consensus
statements.
Estimates of the proportion of the sea that needs to be protected in order
to maintain ecosystem function as it is now, let alone restore it to former
levels, range from 10% (Turpie et al. 2000) to over 50% (Lauck et al.1998).
Consequently, there is a lot at stake and all sides involved are taking
the issue seriously. Ironically, one of the very factors that may make
marine reserves necessary, the difficulty of directly observing the state
of marine environments, also increases the difficulty of establishing the
facts about their level of effectiveness, and what proportion of marine
habitats should be protected. The other reason that factual argument
can be difficult in this emotive debate is that many communities have no
marine reserves of their own. If such reserves do exist, they may be so
new or so poorly studied that it is not clear whether they have been effective
(Jones et al. 1992). In the absence of relevant information, debate surrounding
marine reserves can become polarised and entrenched, more a matter of faith
than anything else, with different groups promoting diametrically opposed
conclusions about their effectiveness (Table 1). Facts can be the first
casualty, and both sides are guilty of making claims that are poorly
substantiated.
One of the main problems polarising those
in favour and against, is familiarity with the sea. Whereas fishermen ply
the sea on a daily basis, doing controlled scientific experiments several
times a day (shooting a net, setting a line), most marine reserves articles
originated from scientists with very little familiarity with the sea or
its ecosystems. It has indeed become fashionable to use computer models
rather than studying the sea. The table below substantiates this. Note
how all myths are on the pro side, the side taken by marine scientists..
Pro-reserve (P-R) reserves will:
Anti-reserve (A-R) reserves will:
protect fished populations [false. Fish are
protected only inside MRs. Migratory fish is unprotected. Most Commercial
fish is migratory and unprotected]
fail to protect fished populations [true. MRs
fail to protect fish outside. They also fail to protect migratory fish.
They fail to protect nearly any fished species]
protect marine habitats [false. Marine habitats
are not threatened. By far most marine species are not threatened. The
effect of MRs on habitat is negligible.]
increase impacts outside reserves [true, but
this argument is unrelated to the one mentioned on left.]
enhance fishing through spillover [false.
It has repeatedly been proved, in NZ and elsewhere that spillover is much
less than the lost fishery]
lock up resources [true. The potential fishery
inside MRs is lost and locked up forever]
export eggs and larvae [false. When large predators
develop, the total output of eggs and larvae from MRs decreases. Recruitment
of commercial fish outside bears no or little relationship to the amounts
of eggs produced inside MRs]
impose extra costs [true. There is the real
cost of the lost fishery, which amounts to hundreds of millions of dollars
in lost export revenue, and there is the additional cost of administration
and enforcement]
provide baselines [false. Not a single MR out
of 1200 worldwide has provided a baseline]
be irrelevant due to existing effective fisheries management system
[true.
MRs do nothing for the fish outside, but fisheries management does. MRs
do not change our ways.]
MRs are urgent.
[false. The situation due to fishing is not getting worse, but the threat
from landbased pollution is. MRs do not protect against this.]
MRs are not urgent.
[true. We must focus on the main threat, land based pollution]
MRs are the only way to protect biodiversity
[false. Fishing does not threaten biodiversity]
MRs do little for biodiversity
[true. Most species are not harmed by fishing. Biodiversity is all about
viable populations of all species.]
Table 1. Examples of apparently contradictory
views promoted by groups either in favour of marine reserves or opposed
to their implementation. These examples derive from the New Zealand
experience, but similar ideas are expressed wherever the issue of marine
reserves is raised.
The above table illustrates clearly how
scientists have not bothered understanding what the 'anti-reserve' skepticists
say. First of all, these are NOT against marine reserves, but they want
them to work and not to fail like the many failed reserves in NZ. These
people do NOT say that marine reserves fail to protect fished populations
inside, but they say that marine reserves do not protect or improve the
fishery outside or prevent it from collapsing or protect against the more
serious risks of mass mortalities, poisonous plankton, oil spills, global
climate change and ozone holes. Marine reserves do not protect the fish
stocks outside but fisheries regulations do!
Typically, Table1 shows on the left all the myths
and on the right all the truths. What makes scientists so ignorant? See
also Frequently Asked Questions in the marine reserves
debate on this web site.
In the Australian context, marine reserves are well represented in some
areas such as the Great Barrier Reef, and evidence of their effectiveness
in protecting exploited tropical species is beginning to emerge (Adams
et al. 2000) [The GBMP was created in 1936. How long
does it take to show benefits?]. However, there is still little
direct experience of marine reserves in much of the rest of the country.
This is particularly true in Australias temperate waters, with the exception
of Tasmania where a range of fish and invertebrate species have been shown
to increase in size and density as a result of protection (Edgar and Barrett
1997,1999). In general, the Tasmanian results show that the effects of
protection are strongest in the largest reserve at Maria Island. Newly
established marine reserves in Victoria are still too young to provide
useful data, and networks of reserves in other parts of Australia are still
in the planning stages. Consequently the New Zealand experience of marine
reserves may provide much needed clarification for the Australian situation,
allowing discussion to be directed in the most constructive directions.
Examples similar to those found in New Zealand are available from around
the world but this paper concentrates on New Zealand because of the many
cultural and ecological similarities between the two countries.
The oldest marine reserve in New Zealand (Cape Rodney to Okakari Pt
or Leigh Marine Reserve) was established in 1976, and New Zealand now has
17 marine reserves spread around much of the country. While some very large
reserves surround remote offshore islands, less than 0.1% of the coastal
zone surrounding the main North and South Island is protected. It is important
to remember that all marine reserves in New Zealand are fully no-take,
with no fishing or extraction of marine organisms. This is not true in
other parts of the world where Marine Parks or Marine Protected Areas (MPAs)
often offer no protection to marine flora or fauna, or protect only a limited
number of species or areas. Typical examples can be found in the Mediterranean,
where Francour et al. (2001) found that amateur and commercial fishing
was allowed in half the MPAs in the Mediterranean, and in Florida, where
99.5% of the Florida Keys Marine Sanctuary provided no protection for any
species (Bohnsack 1997).
Reserve Effects on fished populations Marine reserves are loved by the public. They think it increases
abundancy [sic ] of fish. Science shows it doesn't, but the public believe[s]
it does (Lough 2002).
Despite the fact that marine reserves are now widely recognized as having
positive effects on the abundance and biomass of species within their boundaries
(Roberts and Hawkins 2000), a perception remains in some areas that marine
reserves do not work and that we must still prove they would enhance the
preservation or sustainability of marine species (New Zealand Fishing
Council 2001). There is now abundant information from New Zealand that
can help to clarify this matter.
Scientists concentrated on only a few
exploited species while ignoring the fate of all others. In doing so, they
did not notice the devastating effects of landbased pollution. What use
is a medical checkup when your fatal cancer is not detected? What
use is 'abundant information' when it fails to mention the most devastating
threat to the sea? When will scientists acknowledge that more than two
out of three of our coastal marine reserves are not working?
Spiny Lobster In northeastern New Zealand at the Leigh Marine Reserve (Fig.1; established
1976) the recovery of Spiny Lobster Jasus edwardsii populations
was dramatic, and density increased from around 8 to over 20 J.edwardsii
per 500 m-2 by 1983 (MacDiarmid and Breen 1992). Unfortunately,
no measurements of J.edwardsii density were made outside the reserve
until much later, but they confirmed that the increase in J.edwardsii
was due to a cessation of fishing (Kelly et al. 2000). By 1995 the density
of the J.edwardsii population inside the Leigh Reserve was 3.95
times that of adjacent fished areas (Babcock et al. 1999), while at the
nearby Tawharanui Marine Park (established 1982) the number was approximately
1.6 times greater. At another reserve in the northeast region (Hahei Marine
Reserve established 1992), the number of J.edwardsii was no greater
inside than outside (Kelly 2000). However, the size of J.edwardsii
was greater inside all three reserves.
Look carefully at Fig2 to notice how
the crayfish population suddenly collapsed between 1995 and 2000. The precise
date was June-Oct 1998 when an unusually long period of mud storms drove
the crayfish out. The scientific community was informed of this but did
not want to believe our observations. In fact, two independent groups of
scientists were working with rocklobster while remaining oblivious to a
sudden decrease of 85% in their numbers! Why do scientists repeatedly fail
to mention this in their publications? The Marine Laboratory of the University
of Auckland sits right above the marine reserve and scientists can see
the quality of the water while drinking coffee!
For decades we have been fed the myth that crayfish
densities inside the reserve amounted to over 20 times that outside, yet
the above-mentioned factor of 4 is more realistic. Why have scientists
been misinforming the public for so long? Recent publications now mention
a factor of 1.6, commensurate with the situation after 1998. The main
message is that marine reserves do not make a major difference and
marine
reserves do not protect against degradation.
Based on these three marine reserves of differing ages, an average rate
of increase of 7.4%yr-1 was estimated (Kelly et al. 2000). The
history of recovery at Leigh suggests this would be a minimum (Fig.2) with
numbers increasing by over 4.5 times between 1978 and 1983 (MacDiarmid
and Breen 1992). In parts of the country, where J.edwardsii recruitment
is high, such as along the east coast of the North Island, the rate of
increase has been even higher, increasing by 6 times after just 4 years
at Te Angiangi marine reserve (established 1997, Freeman and Duffy in prep).
In the Tonga Island marine reserve (established 1993) in the northern South
Island,
J.edwardsii has also shown significant recovery (Davidson
et al. 2002) but not all reserves have shown these increases. The Te Awaatu
marine reserve (est. 1993) in Fiordland had not shown a significant increase
in
J.edwardsii numbers after 6 years of protection (Kelly 1999).
Whether this was due to its size, the nature of the fiordland habitat,
seasonal factors or the continuous decline of the surrounding fishery (Starr
et al. 1997) is not clear. Certainly the North Island regions support stable
fisheries.
The quoted crayfish recovery rate of
7.4% per year is another myth. Watch Fig2 for the course of a typical recovery,
which levels out after a decade. This is for the Goat Island marine reserve
which is endowed with an unusual amount of good crayfish habitat. The first
decade saw 400% in 10 years, followed by 0% in the next ten years, and
a collapse of 85% in one year. Where does that leave the quoted 7.4% per
year? The recovery rate is not only dependent on the quality of the habitat
but also on the state of the fishery. Generalizations are dangerous.
Fig.1. Map of New Zealand featuring key areas and marine
reserves mentioned in the text. Detail shows northeastern New Zealand which
has the highest density of reserves and where most of the work on marine
reserves has been conducted. Not all reserves are marked on this map.
Fig.2.
Recovery of spiny lobster population in the Leigh marine reserve. While
no data were collected prior to protection and were only sporadic until
the mid 1990s the increase in density is most likely due to protection
and has been observed in other reserves around the North Island. The reason
for a substantial decrease in numbers at the Leigh reserve since 1995 is
not known but has not been observed at other reserves (e.g. Hahei). Data:
Ayling 1978, McDiarmid and Breen 1992, Kelly et al. 1996, Kelly and Haggitt
2000. Triangle= time of reserve creation (1976).
The sudden decline of rocklobster
was caused by prolonged mud storms between June and September 1998, which
caused a massive walk-out. It is amazing and unforgivable that scientists
did not notice the event and its consequences. The rock lobsters were all
caught in the same year, outside the reserve. Divers and fishermen experienced
record catches from the area around Leigh Harbour.
Snapper The restoration of snapper Pagrus auratus populations in marine
reserves is now well documented in northeastern New Zealand. This species
proved difficult for divers to census (Cole 1994) and it was not until
Baited Underwater Video (BUV) census methods were employed (Willis et al.
2000) that the magnitude of their recovery could be measured (Fig.3).
The BUV is one of the worst measuring
devices ever invented since it conflicts with basic principles. All measuring
instruments are designed such that they cause the least influence on the
quantity measured. By contrast, the BUV maximises its influence by attracting
fish from afar by smell and by offering food. Such an instrument will exaggerate
the quantity measured. Its linearity or one-to-one correspondence between
observed and actual quantities has not been proved. Scientists have not
taken the necessary precautions with the results produced with this device,
results which are way out of line with those found elsewhere. The method
is sensitive to the currents and hunger level of fish.
A comparison of three coastal Marine reserves showed an average ratio of
14.3 times more P. auratus above the legal minimum length (270 mm)
inside as opposed to outside reserves (Willis et al. 2003). For all fish,
including juveniles, the reserve:Non-Reserve (R:NR) ratio varied, being
4.2 at Leigh, 2.4 at Hahei, and 2.1 at Tawharanui (not significant). The
R:NR ratio for legal sized P.auratus was similar among the three
reserves at the time of the study, even though the youngest reserve was
6 years old and the oldest was 23 years old. Snapper recovery therefore
seems to be rapid, an impression confirmed by the increase in P.auratus
numbers at the Poor Knights Islands Marine Reserve.
Scientists hail these figures without
thinking about their consequences. Having 14 times more fishable snapper
in a closed area means that the fishery outside teeters at less than 7%
(5%?) of unexploited stock, which is not borne out by fisheries statistics.
Obviously, the measuring method is seriously flawed.
Fig3 shows how Goat Island stands out above the
rest. Scientists fail to mention that Goat Island has an unusual amount
of reef habitat and shelter, and that the centre of the reserve is located
around an island. Fish congregate here because it offers clean water (much
cleaner than 400m further inshore), currents, deep habitat close to feeding
grounds and ample shelter from storms. It is an unusual place. It so happens
that the other two places, Hahei and Tawharanui were also chosen because
of their good qualities for diving. They too are not representative of
the rest of the coast. Why do scientists fail to mention this?
The Poor Knights Islands were declared a partial marine reserve in 1981.
A small proportion of the area was no-take but recreational fishing, using
unweighted lines, was permitted in the majority of the reserve. In October
1998 the entire reserve reverted to no-take. At that time there was a
significant [just measurable] difference in
legal P.auratus abundance between fully no-take areas of the Poor
Knights and those areas with limited fishing, this difference was small
in magnitude and likely to have been of limited ecological significance
(Denny et al. 2002). After 3 years the number of legal-sized P.auratus
had increased by 8.3 times relative to the pre-closure values (Fig.4) and
were 16.6 times more abundant than in fished reference areas, where numbers
of legal sized P.auratus have remained static (Denny et al. 2002).
Snapper populations at the Poor Knights Islands demonstrate that while
rapid recovery is possible, it is only likely to occur under fully protected,
no-take conditions.
Fig4 shows how quick a fishery recovers
after total closure, which basically says that the ecological effect of
fishing is a minor issue and so are the claimed benefits of reserves. What
scientists fail to disclose is the unique situation of the Poor Knights
Islands. These are located at the edge of the continental shelf, surrounded
by 80-120m deep sea bottom. Snapper from a wide area around migrate on
a regular basis to the only shallow warm water nearby. Thus measuring snapper
densities in shallow water distorts reality and is not representative of
densities in the surrounding deep sea bottom, the main extent of this reserve.
To compare this oasis sanctuary with the Mokohinau Islands, is rather naive.
Again no further thought is paid to the consequences of the factor 16,
which implies that fish stocks elsewhere are below 5% of unexploited levels.
Why are marine scientists so ignorant and naive, one may ask.
Similarly, the Mimiwhangata Marine Park (established 1984) on the coast
adjacent to the Poor Knights, allows only recreational spearing or fishing
using unweighted lines. Despite the exclusion of commercial fishing, the
abundance of legal P.auratus at Mimiwhangata was not significantly
different to reference areas outside the park. In fact, mean P.auratus
numbers there were lower than at any other areas, despite the recreational
gear limitations and the complete absence of commercial fishing (Denny
and Babcock 2002).
The Mimiwhangata Marine Park extends
outward by only 100m. It is dominated by sandy beaches and much shallow
sandy seabottom. There is little reef habitat. It is not policed and most
people fishing there do not know that fishing rules apply.
The magnitude and speed of recovery of P.auratus populations in
northeastern New Zealand can probably be ascribed to two factors. Firstly,
individuals of this species can display a variety of behaviours, including
seasonal onshore and offshore movements (Willis et al. 2003), as well as
long-term residency within restricted areas (Willis et al. 2001). It is
also likely that individuals switch from one behaviour to the other, and
that they show intermediate types of behaviour. The seasonal migratory
behaviour means that densities of legal P.auratus inside reserves
vary by 3.9 times between spring (September) and autumn
(April) as fish move on and offshore from coastal reefs (Fig.3, Willis
et al.2003). A proportion of these fish take up residence on the reefs
where they may remain in home range areas of less than 300 m diameter (Parsons
et al. 2000) for periods of up to 3 years (Willis et al. 2001). In one
case, a tagged P.auratus was seen 6 years after tagging, at less
than 1 km from the tagging site (R. Babcock, unpublished data). Newly created
reserves are therefore quickly stocked with migrating fish, some of which
are likely to take up long term residence. The second reason for the ability
of P. auratus populations to recover is that there is a substantial
stock of fish in northeastern New Zealand which provides not only migratory
fish but also larval recruits (Gilbert et al. 2000).
This is indeed what fishermen are seeing.
The Leigh marine reserve has some large resident snapper (e.g. Mister Perfect,
Panda and Monkeyface) which disappear between November and March to spawn
outside the reserve. Miraculously they dodge fishing lines and nets, to
reappear in April/May. This suggests that stocks of large old snapper exist
outside marine reserves, experienced enough not to take the bait. The main
message here is that marine reserves do little for migrating stocks, which
make up the bulk of commercial fishing.
Reserves in other areas that once supported significant P.auratus
populations, e.g.Tonga Island marine reserve in the northern South Island,
do not appear to have shown a marked recovery in P.auratus populations
(Cole et al. in press). The region once supported a substantial P. auratus
fishery but this has been in serious decline since 1979 when the annual
catch peaked at around 3203 tons and it now less than 200t (Harley and
Gilbert 2000). The yield of the fishery in the Hauraki Gulf (northeastern
New Zealand) has remained at over 5000 tons per annum since the 1940s.
In the case of both P.auratus and J.edwardsii, the restoration
of populations in marine reserves appears to have been facilitated by the
presence of adjacent healthy populations.
This is precisely what fishermen are
saying. Marine reserves have very little benefit to commercial and recreational
fishing. They do NOT protect fish stocks. It is much better to manage fish
stocks at higher levels everywhere than to hope that locking up small areas
will help.
Fig.3. Relative abundance of Snapper (Pagrus auratus)
inside
and outside three marine reserves in northeastern New Zealand. Data are
for fish greater than minimum legal size (270 mm) from 30 min deployments
of Baited Underwater Video (BUV). Filled symbols: marine reserves, Open
symbols: adjacent fished areas. Leigh, Hahei, Tawharanui. (after Willis
et al.2003). Circles= Leigh; triangles= Hahei; squares= Tawharanui.
1) the BUV exaggerates the observations.
2) there exists a large difference between spring and autumn, which could
mean that fish move away or that they are more or less well fed. 3) the
results from Goat Island swamp the rest because it is a special place.
4) these three reserves are all more special than the areas outside.
Fig.4. Increase in relative abundance of Snapper population
at the Poor Knights Marine Reserve relative to fished Reference areas.
The entire Poor Knights Island Group was declared a no-take reserve in
October 1998. Data are for fish greater than minimum legal size (270 mm)
from 30 min deployments of Baited Underwater Video (BUV). Circles= Poor
Knights; triangles= Cape Brett; squares= Mokohinaus. (after Denny et al.
2002). Triangle= time of reserve creation.
1) The Poor Knights are an oasis
type of reserve. 2) the amount of shallow warm water is small compared
to the dominant deep sea bottom. 3) fish migrate regularly from deep
to shallow water, thus concentrating in the shallows, which distorts the
data. 4) the BUV exaggerates. 5) recovery is fast, suggesting that the
detrimental effect of fishing is small. 6) Marine reserves have little
to offer. 7) controlled fishing did not affect the rest of the reserve
or the enjoyment of diving there. 8) A rigidly enforced marine reserve
is not better than a voluntary one.
These two graphs (top Baited Underwater
Video and bottom Visual Underwater Census) are from the same study as of
Fig.4, showing the total snapper count for Poor Knights (circles), Mokohinau
(triangles) and Cape Brett (open squares). Scientists should have drawn
regression curves (red curves) rather than comparing points 1 and 7, as
they have missed the high point of 1998. However, as can be shown, the
difference between Poor Knights and Mokohinau is almost negligible, which
refutes these scientists' claim.
Please note that the bottom graph
was left out of the report. Why?
The data in this report also showed
chronic decline of the common coastal fish species which belong to the
Poor Knights and which breed there, but not a word of this phenomenon can
be found in the report.
As can be seen, nearly all species
suffered two to six-fold declines, except for sweep which have been increasing
their numbers. Sweep compete with blue maomao for space but they belong
to the more turbid waters along the coast. Their increase at the Poor Knights
means that turbid waters have arrived there while at the same time numbers
of blue maomao have declined, which is what we have observed.
The scientists who conducted this
study have arbitrarily decided not to count schooling fish, so the sudden
decline of blue maomao, trevally, koheru, jack mackerel, kahawai and to
some extent also pink maomao, has not been recorded. One may ask how relevant
such studies are when they are either incomplete or not reported properly.
This is a relevant question since taxpayers paid $280,000 for this four-year
study.
To put it all in perspective, children
are now learning at school that snapper increased 16-fold after fishing
was stopped at the Poor Knights!
Direct Effects on marine ecosystems: habitat recovery in taking an approach that wants to protect biodiversity, we'll
also ensure that fish stocks are there and the bio-mass is maintained
(S.Lee. The Dominion ,7 February 2002)
The effects of fishing at the ecosystem level are now evident worldwide
and extend far beyond the target species (Pauly et al.1998). The direct
effects of fishing, mainly trawling and dredging, on marine habitats are
also widespread and pervasive and have been estimated to disturb areas
of sea floor equivalent to the world s continental shelf once every two
years (Watling and Norse 1998). Three dimensional habitat structure is
reduced or destroyed by these forms of fishing but no-take MPAs in locations
such as the Georges Bank (northwest Atlantic) have enabled recovery of
this habitat structure to occur (Collie et al. 1997).
To quote the above references is more
than just malicious, particularly with respect to New Zealand, where fishery
management is making vast improvements. To quote that trawling disturbs
the sea floor once every two years is ignoring the fact that storms do
so several times each year. It is also showing ignorance of the methods
of trawling, some of which cause very little disturbance.
Three dimensional habitat structures are often
quoted, but do these really exist on a sandy and muddy bottom, which is
the vast part of the continental shelf? Sponges and gorgoneans are associated
with rocky substrate, which is what trawlers avoid. Only occasionally do
trawlers bring up sponges living on low-profile rock flats in areas of
current. Such places are ephemeral, often smothered by sand. Most likely
these organisms are just opportunistic but unimportant.
In NZ we have vast de-facto marine reserves on
the seabed in a continuous range of depths, yet no research has been done
on these. No plans have been made to convert these to marine reserves.
Why do scientists avoid these? What is their political motivation?
Species assemblage and habitat structure of soft bottom communities in
northeastern New Zealand have also been linked to the intensity of trawling
and dredging, using marine reserves as reference areas (Thrush et al. 1995,
1998). On Australias northwest shelf, a ban on trawling has allowed sessile
macrobenthos to recover. The increase in habitat-forming species, such
as gorgonians and ascidians, has been paralleled by an increase in the
numbers of target fish species associated with them, such as tropical snappers
and emperors (Lutjanidae, Lethrinidae; Sainsbury et al. 1998). Whether
this was a direct result of the cessation of fishing, or an indirect effect
of habitat structure, remains unclear [after all
the above statements?]. Only long-term and large-scale experiments
will tell us conclusively whether changes in habitat structure affect the
productivity of commercial species.
Ecological principles say that habitat
structure on soft benthos is just opportunistic and ephemeral (short-lived)
and of no importance to the environment.
Displacement of fishing effort More marine reserves could force fishers into the remaining areas,
depleting stock (A.Macfarlane, New Zealand Herald, 4 February 2002.)
Some attempts to model marine reserves have concluded that no-take areas
as large as 40-80% of the available habitat may be required in order to
sustain fisheries yields (Sladek Nowlis and Roberts 1998), or ecosystem
integrity (Sala et al. 2002). Other models show that if fishing effort
remains constant while available fished area decreases the displacement
of fishing effort by no-take areas could increase the impact of fishing
on unprotected areas (Parrish 1999). Thus while some areas would be less
impacted by fishing, the effects would increase proportionally in other
areas.
Marine Reserve scientists, in their zealous
ideology, have attempted to use marine reserves as a fisheries management
tool. In tropical seas where artisanal fishing cannot easily be controlled,
such may be possible, although no good examples exist so far. Using computer
models, these scientists have let their fantasies loose on a wide range
of scenarios, all of which weakened by their underlying assumptions. The
underlying problem is that a good reserve which fosters wise old fish,
does not leak fish easily to the outside. But it closes the area to fishing.
Thus the benefit to fishers is negative. Spill-over does not compensate
for the lost fishery. If 20% of the sea is closed off, the public loses
20% of its fishery, which converts to $200M in lost exports every year.
Is this our legacy to our children? For more about the 10-80% craze, read
Target
sizes for marine reserves.
The problems foreshadowed by this model are of genuine concern, but their
potential importance depends on several factors. Firstly, the proportion
of coast set aside as marine reserve would have to be much larger than
is seriously being proposed anywhere at present. In New Zealand, target
proportions advocated by pro-reserve groups range from 10 -20%, varying
from one interest-group or political party to the next. At these levels
of protection, the amount of effort displaced to other areas of coast will
be relatively small (Fig.5). The second condition would be the absence
of effective fisheries management regimes outside protected areas. Proposals
for systematic large-scale networks of marine reserves do not include dismantling
existing management systems.
Indeed, fishermen go one step further
by saying that marine reserves are not necessary for achieving sustainable
fishing. They may be necessary for research, education and diving, but
not for fisheries management. What's more, so far not one out of 1200 MPAs
worldwide, including those in NZ, have proved to be of benefit to fisheries
management but a temporary closure such as at the Georges Bank will work
as a normal fishery management tool.
In the New Zealand context, a figure of 20% representation of coastal areas
in MPAs is the highest proportion currently being proposed (Green Party
2002 and Forest&Bird) and the Labour government
supports a total of 10% (Department of Conservation 2002). This figure
is well below that at which we would expect to see a rapid increase in
displacement to unprotected areas. Furthermore, the current New Zealand
Fisheries Act (1996) requires that fisheries are managed to ensure the
protection of fisheries habitat as well as fished species. Consequently,
fisheries management of non-reserve areas will need to account for and
mitigate against undue pressure on fisheries and habitats as a result of
any fishing displacement. The greater the proportion of marine reserve,
the greater the need for managing human uses of non-reserve areas. To paraphrase
the title of a frequently cited paper (Allison et al.1998), while fisheries
managers may not agree that marine reserves are necessary for marine conservation,
they understand well that reserves alone will not be sufficient. Therefore,
to raise the issue of displacement of fisheries effort as a major drawback
of marine reserves requires us to adopt some unrealistic positions. This
would include a simultaneous enhancement of management and conservation
efforts within marine reserves, and a relaxation of these efforts in fisheries
management. [oops, a non-sequitur]
The consequence is that marine reserves
should be managed under the Fisheries Act while the Marine Reserves Act
should be abolished. This would also resolve conflicts imposed by one department
on another, and of one part of society on another. Marine reserves should
be created for what they do best: research, education, diving.
Fig.5. Exploitation rate in fished areas as a function
of the proportion of the fished stock in marine reserves. The model assumes
that total catch and effort remains constant and that the fished stock
is evenly spread around all areas of coast. If this is so, the proportion
of protected stock is proportional to the protected coastline, and in order
to maintain catches, effort must go up in the remaining fished areas. (After
Parrish 1999).
This graph is in fact, quite meaningless.
It compares two incomparables, exploitation rate and biomass rather than
reserve size. It assumes that catch and effort remain constant even though
exploitation rate goes up. What the public knows intuitively, is that if
you lock up 20% of the fishery, the remaining 80% of the sea cannot be
fished more intensively.
Indirect Effects on marine habitats: environmental
baselines Certainly they are great for science, to study the sea and the
relationships between a range of animals on a long term basis (J.Nicolson,
New Zealand Herald ,31 January 2002).
Because of the pervasiveness and intensity of fishing activities, it
has been suggested that marine reserves offer virtually the only way of
understanding what a natural ecosystem might be like, or of appreciating
the full impacts of fishing (Dayton et al.1998). In addition to the physical
damage caused by fishing, one obvious reason for this is that fishing may
have directly reduced populations of some species to the point where they
are no longer functionally-significant ecosystem components.
Dayton talks about environmental changes
having happened so long ago that nobody can remember what the world (or
sea) looked like. How frighteningly true. But he overlooks that the sea
has not suffered the irreversible damage of forest logging and burning,
agriculture and urbanisation. By comparison, it is still a pristine environment.
No doubt, fishing will have left its mark but this can in no way be compared
to what humans have done on land. Recovery in the sea happens in a matter
of one decade compared with centuries for a forest. What all scientists
seem to overlook is that the damage from landbased pollution has caused
permanent and long-lasting change to the marine environment. Fishermen
are now saying that over 40% of the coast anywhere is no longer suitable
for fishing. Yet scientists choose to ignore this.
People who blame pervasiveness and intensity
of fishing, simply demonstrate that they have little understanding of marine
ecology and its energy flows. It is sad.
Indirect effects of fishing on marine habitats are less obvious but are
no less important [?]. Marine reserves in
northeastern New Zealand have provided important examples of such indirect
effects and the importance of reserves as environmental baselines. The
first quantitative ecological descriptions of rocky reefs on the northeastern
coast of New Zealand described extensive areas of urchin barrens, largely
devoid of macroalgae and dominated by the grazing echinoid Evechinus
chloroticus (Ayling 1978, Choat and Schiel 1982). Such barrens areas
were extensive in no-take reserves, such as those at Leigh and Tawharanui,
at the time of their establishment (1976 and 1982 respectively).
By the 1990s, the extent of urchin barrens areas at Leigh was much
smaller than it had been in the 1970s [this happened
suddenly in 1995-98 and not gradually as is suggested], and at both
Leigh and Tawharanui reserves the proportion of reef occupied by barrens
habitat was significantly lower inside than outside reserves (Babcock et
al. 1999). Based on these comparisons, it was estimated that primary productivity
of reefs might be as much as 58% greater on reefs inside the reserve than
outside, due to the increase in biomass of brown algae such as Ecklonia
radiata (Babcock et al. 1999). Densities of P.auratus and J.edwardsii
inside both reserves were correspondingly higher than in fished areas,
therefore one indirect effect of fishing was a trophic cascade in which
densities of these predators controlled urchin densities and, indirectly,
algal biomass and productivity (Fig.6). Based on more conventional small-scale
manipulative experiments (Andrew and Choat 1982, Andrew and MacDiarmid
1991, Steinberg et al. 1995), such a trophic cascade was thought not to
exist in New Zealand.
[Sigh] When does the author realise that
the disappearance of the urchin barren zone was caused not by the beneficial
effect of a marine reserve but by degradation (kelpbed death Jan 1993,
urchin deaths 1993-2002, crayfish walk-out 1998)? For a complete rebuttal
of these untenable claims, see Science Exposed.
Urchin population structure and behaviour within the Leigh and Tawharanui
reserves differed from that found in adjacent fished areas (Cole and Keuskamp
1998, Shears and Babcock 2002, 2003). More urchins adopted cryptic behaviour
within reserves, sheltering among or beneath boulders. This was true even
at sizes above the threshold (35-45 mm test diameter) at which they begin
to graze openly on the substratum in fished areas. Populations in reserves
tended to be bi-modal, with 35-45 mm size classes poorly represented, presumably
due to predation.
The author and other marine scientists
do not sufficiently make a distinction between the exposed north facing
coast of Goat Island and the more sheltered south-east facing coast which
is used for comparison. Urchins on exposed coasts stay cryptic (hidden)
longer, due to wave action. This can also be seen in other places. The
bi-modal (two peaks) population says that one or more year classes are
missing due to mass mortality.
Tethering experiments showed that small urchins were most vulnerable to
predation and confirmed that predation was higher inside reserves and that
at least 45% of urchin mortality was attributable to J.edwardsii
(Shears and Babcock 2002). One consequence of changes in urchin behaviour
and size-specific predation may be a significant time lag in the manifestation
of trophic cascade effects. Decreases in urchin density and habitat transitions
from barrens to kelp or algal turf were still being recorded in the Leigh
reserve as recently as 2000 (Shears and Babcock 2003). These changes in
habitat may in turn have indirect effects on the abundance of other organisms.
For example, the density of the limpet Cellana stellifera is lower
inside reserves while the density of the turban shell Cookia sulcata
is higher (Shears and Babcock 2003). Experimental habitat manipulations
have shown that Cellana grows and survives better in the presence
of Evechinus, while the reverse is true of Cookia (Andrew
and Choat 1982).
Marine reserves in New Zealand have demonstrated their potential to
act as environmental baselines and ecological tools, and have enabled insights
to be made into the management, productivity and ecological function of
coastal ecosystems that would not otherwise have been possible. More insights
are likely to be obtained from the range of more recently created marine
reserves in other parts of the New Zealand, as well as Australia. These
may take some time to appear if timelags, such as those seen for trophic
cascade effects in northeastern New Zealand, are involved. Systems in areas
where urchin barrens are present, such as the northern South Island, and
the NSW coast (Andrew and O Neill 2000), are likely to respond with trophic
cascades similar to those seen at Leigh and Tawharanui. However, there
are indications that not all systems will respond in this way. For example
macroalgae, not urchin barrens, dominate habitats around the majority of
the New Zealand coastline (Choat and Schiel 1982) and much of temperate
Australia (Prince 1995, Fowler Walker and Connell 2002, Edgar and Barrett
1997).
By being so keen to demonstrate 'effects'
of marine reserves, marine researchers have missed the opportunity to learn
more of the marine environment. For instance, our own observations as measured
in Survey93, have largely
been overlooked and ignored by scientists, even though these findings are
critical to understanding the marine ecology. Science these days appears
to consist of snapshots to prove an idea right, rather than following a
sequence of ideas and experiments to understand how things work in the
sea. Because of this, the author has missed what was really happening in
the environments he studied. The fact that most of the research was funded
by the Department of Conservation, did not help either. How much trust
would you place in health research funded by the tobacco industry?
Fig.6. Habitat change at Leigh Marine Reserve. Trophic cascades
resulting from the recovery of predator populations after
fishing ceased led to a transition from urchin barrens
dominated by Evechinus chloroticus (a) to mixed algal assemblages
(b). Both photos are of the same site, at 5m, facing
northwest from Martins Rock in 1993 (a), 2000 (b).
Again, no mention is made of the
mass kelpbed die-off in 1993, mass urchin deaths and the crayfish walk-out
of 1998.
Effects on fishing: lock-up or spillover? the international evidence of increases of stocks in fisheries
next to established marine reserves remains indisputable (S. Lee,
The Dominion, 7 February 2002)
What I am condemning is the objective of locking up a vast amount
of perfectly good ocean in marine reserves for all time The kaftan wearers
may shout what about spillover , but paua [abalone ] don t spillover
(E.Arron,
Dive NZ June/July 2002)
If the government is successful in locking up 10 percent of New
Zealands fishing grounds there will be significant implications for commercial,
customary [maori ] and recreational fishers. (N.Gibbs, New
Zealand Herald , 31 January, 2002)
No other concept associated with marine reserves is more controversial
than that of spillover. Spillover implies that fish may move out of a
protected area due to density-dependent effects, once the area approaches
carrying capacity (Kramer and Chapman 1999). The use of this term has broadened
to include any movement across a reserve boundary. Since reserves do not
physically fence in fish, cross boundary movements are inevitable. Unfortunately,
little is known about the scale of these movements.
The author fails to mention the concerted
effort made by many scientists to measure spillover, all to no avail. It
simply is not there or is too little to be of value.
Because the opposing claims of those who support or oppose marine reserves
hinges on this knowledge, many apparently contradictory claims can be found
in the scientific literature as well as in popular media. Fishing interests
generally hold the position that reserves will lock up fish resources,
implying that fish will not cross reserve boundaries and that they will
not be available to the fishery (Te Ohu Kai Moana 2001). This is often
accompanied by claims that reserves will not protect fish populations because
fish move too widely (Option4 2002).
This is also supported by science. Fish
will move across boundaries, but not noticeably more out than in. Besides,
a good reserve does not want to lose its breeding stock. The point here
is that the fish species of commercial interest, roam widely and in high
numbers over vast stretches of sea bottom or open sea. In order for fish
to benefit, they must remain inside the reserve (be site-specific) and
also of interest to fishers. Red Moki (Cheilodactylus spectabilis)
is such a fish although it is not keen to take the bait but it is of little
commercial interest due to its low numbers. Note in this respect that for
fish to be commercially attractive, they must occur in large numbers, which
requires large habitats like the flat sea bottom. On such monotonous habitats
the fish do not stake out fixed territories, so they all roam about. The
rocky shore habitat where fish are more resident, is too small to provide
for enough commercially attractive fish.
Proponents of marine reserves claim that cross boundary movements of fish
(presumably combined with a longer period of growth while protected) will
result in enhanced fisheries yields at some scale (Roberts et al. 2001).
Given
the diversity of fish species and their diverse life histories, generalisations
are unwise, but this does not seem to have tempered the statements of either
side [!]. While little evidence exists to
support either claim, the facts that do exist tend to support the idea
that reserves will enhance yields (Russ and Alcala 1996, Roberts et al.
2001). Even so, much of this evidence has been questioned because of concerns
about replication or other design aspects of the studies (Hillborn 2002).
Marine reserve skeptics, please note!
Finally a confession and another myth can be laid to rest, or will it?
Evidence from New Zealand indicates that while reserves do not lock up
resources [oops], neither do they enhance
fisheries yields (Fig.7). Rather it seems that, as far as can be measured,
the effects of reserves on local fisheries yields are neutral [oops,
what about the lost fishery within?]. A three-year study of a lobster
fishery in the Leigh region compared Catch Per Unit Effort (CPUE) and value
of catch around the boundary of the Leigh marine reserve with those of
two other areas in the region, Coastal Leigh and Little Barrier Island.
Overall, both CPUE and value of catch were the same around the Leigh reserve
boundary as they were at the two other areas (Kelly et al. 2002). This
was an unexpected result since fishing adjacent to the reserve took place
at the seaward boundary, 800m offshore. This is an area of sandy bottom
up to 700 m from the coastal reefs where J.edwardsii makes its dens,
and where fishers usually set their traps.
The author fails to mention that the
marine reserve displaced crayfishermen who were then forced to place their
traps at the reserve's boundaries, left, right and outward. It did not
take long for them to discover that such was a waste of time, and they
gradually moved to places further away.
Fishing along the offshore reserve boundary began around 1985, when the
reserve was nine years old. Fishers target the offshore boundaries of the
reserve during the seasonal movements of J.edwardsii when they travel
away from reefs onto the adjacent sand flats. The reasons for these movements
are not entirely clear but they are associated with periods of increased
feeding activity (Kelly et al. 1999). Jasus edwardsii can travel
for several kilometers during these movements that may last for weeks.
When Spiny Lobster return to the reef it is usually to the same part of
the coastal reef they occupied previously and some individuals have been
tracked back to the same den at which they were originally tagged (Kelly
2001). This combination of site fidelity and movement over scales similar
to the dimensions of the reserve has resulted in a situation in which J.edwardsii
populations are afforded a substantial degree of protection while still
making a significant contribution to local commercial catches.
The outside boundaries have always been
targeted, since the beginning of the marine reserve. Trawlers discovered
that they could take substantial 'cashies' (crayfish for cash on the black
market) in a single swoop, however, to the detriment of local craypots
which also disappeared in the nets. See Lessons from
Leigh for more details.
The marine reserve at Leigh has protected the resident J.edwardsii
population, but it has not locked it up, contrary to claims of some reserve
opponents. However, the study of the J.edwardsii fishery at Leigh
has shown that one of the claims of marine reserve advocates, that marine
reserves increase yields, may be exaggerated. Yields of J.edwardsii
are
no higher adjacent to the reserve than they are at other comparable sites
in the region. Overall, the fact that a positive conservation outcome has
been achieved without detriment to the commercial fishery would seem to
be a desirable result. [oops]
When the crayfish walked out en-masse
in 1998, the cray fishers said "The reserve is finally working because
the crayfish finally spill out". They had a good year and that was
that.
This author is consistently wrong about 'no
detriment to the commercial fishery'. What about the lost fishery inside
the reserve?
Fig.7. Lobster catch around Leigh Marine Reserve. CPUE
from three years of fishing on the offshore boundary of the Leigh Marine
Reserve and on adjacent coastlines. CPUE at each site varied from year
to year but overall did not differ among the sites adjacent to the reserve
and non-reserve sites. (after Kelly et al. 2002). White= Leigh reserve
boundary; black= coastal Leigh; shaded= Little Barrier Island.
These measurements show clearly
that there is no measurable spillover from the Leigh marine reserve. It
is a pity that the series was discontinued one year before the massive
crayfish walkout in 1998, which demonstrates yet again that scientists
are not interested in how things work but only in scoring political points.
It is sad.
While no direct measures have been made of the influence of marine reserves
on catches of New Zealand fish species, at least one of them, P.auratus,
has characteristics similar to J. edwardsii that may result in a
similar balance of conservation combined with a contribution to fisheries.
[oops]
A high level of site-fidelity is shown by some snapper, with home ranges
in the order of less than 300 m radius as demonstrated using visible and
acoustic tags (O Dor et al. 2001, Willis et al. 2001). A proportion of
individuals show a wider ranging behaviour, and acoustic tracking in the
Leigh reserve has shown that they may range over distances of more than
1 km (Egli and Babcock 2002).
Some of these tracked fish have left the reserve for periods of up
to several weeks before returning. This behaviour, combined with the seasonal
movement patterns of P.auratus (Crossland 1976), indicates that
in addition to the conservation benefits derived from reserves [?],
there is a clear potential for fish that have spent some time in a reserve
to re-enter the fishery. Similar conclusions have been drawn for Blue Cod
Parapercis
colias based on a mark-recapture study in central New Zealand (Cole
et al. 2000).
School children visiting the Leigh marine
reserve enjoy the presence of some large resident snapper. They keep their
fingers crossed that these legendary fish will NOT re-enter the fishery.
But a reserve offers little protection.
A spatially explicit model of the Hauraki Gulf Snapper fishery, incorporating
movements of both fish and fishers, suggests that even with 50% of the
area set aside in no-take areas, there would be virtually no impact on
the yield of the fishery. Under the various scenarios explored, the influence
of no-take areas on catch rates might be either positive or negative (Bentley
et al. unpublished), but the magnitude of any effects was predicted to
be small in relation to the overall catch. This is in agreement with the
empirically derived conclusions of Kelly et al. (2002) in their small-scale
study of the J.edwardsii fishery around Leigh.
Don't believe it. These models are based
on untested assumptions. One needs to be naive to be impressed. See myths(6)
for more admissions from marine scientists.
Egg and larval export marine reserves will boost fishers catch rates by giving fish
a safe place to spawn and rebuild their flagging numbers (S.
Lee The Dominion ,7 February 2002)
The potential for marine reserves to contribute disproportionately large
amounts of egg production is an idea widely promoted as a way that marine
reserves can benefit the stock outside of reserves (Roberts 1997). Since
eggs and ultimately larvae are likely to be transported out of any reserve,
even though some may be retained (Jones et al. 1999), protected populations
in reserves may help maintain recruitment at larger scales. There is clear
evidence that areas of coastline protecting populations of commercially
exploited species do indeed contribute disproportionately to egg production.
Because both numbers and size of protected species increase because of
protection, there is also an increase in the biomass of protected species,
and egg production is proportional to biomass.
We are now entering the vagaries of egg
and larval production, so keep in mind that a mature snapper produces many
millions of eggs in order to reproduce itself perhaps once a year. So most
of these eggs are wasted. Why?
The energy of the sun is packaged into tiny plants
in the plant plankton, which are of no use to larger organisms. The whole
purpose of spawn overkill is to produce food rather than offspring. Larvae
of all sizes eat the phytoplankton and each other, resulting in ever larger
food packages. In the end, snapper live indirectly from their own offspring.
It is a method although inefficient for each, is indispensable for all.
The bottom line is that egg production and reproduction are almost unrelated.
So a successful spawning year bears no relationship to the size of the
spawning stock or the number of eggs produced. This has also been confirmed
scientifically (read the Shipp report).
As can be expected, no scientific evidence has been found for protected
stocks benefiting larval recruitment.
For P.auratus, egg production inside reserves is estimated to be
18 times greater inside than outside reserves, based on a study of three
reserves over three years (Willis et al.2003). [oops,
Fig3] Similar results have been demonstrated for J.edwardsii
in northeastern New Zealand, with egg production increasing at 6.7% per
year (Kelly et al. 2000). After 25 years, this rate of increase would equate
to egg production 4.4 times greater inside than outside reserves (Kelly
et al. 2000). [oops] In the case of P.auratus,
these differences mean that a reserve covering approximately 5 km of coastline
(similar to the marine reserve at Leigh) would produce a number of eggs
or larvae equivalent to 90 km of coastline (Willis et al.2003) [oops].
For J.edwardsii, the equivalent length of coast could be from 22
km (Kelly et al. 2000) to 80 km (MacDiarmid and Breen 1992) depending on
when the estimate was made. In principle, the relatively small no-take
reserves have the potential to sustain recruitment in much larger portions
of the coast.
Here we have landed in the middle of
unsubstantiated assertions where myths are born. 1) the BUV exaggerates
fish counts. 2) the three reserves are rather special. 3) there is no evidence
that snapper populations are increasing indefinitely. 4) extrapolation
over 25 years is mischievous. 5) extrapolation over the entire coast, including
sandy shores, is equally mischievous. 6) what happened to the previously
stated presence of adjacent healthy populations? 7) the total amount
of spawn mass from reserves is smaller because there is less from prey
species.
Stock-recruitment relationships in fished species are notoriously weak.
Some fisheries scientists argue that, because of this, it is unlikely that
any contribution from reserves to overall recruitment would be undetectable
against the background of environmentally determined recruitment variability.
Indeed, variations in seawater temperature explain 94% of annual recruitment
variation for P.auratus in the Hauraki Gulf (Francis 1993). One
of the largest Marine Protected Areas in US waters covers 17,162 km2
(<30%) of the Georges Banks and it has achieved a marked recovery of
scallop stocks (Murawski et al. 2000). Scallop biomass is 9 times greater
in the closed areas than in trawled areas [not insignificant!
but disputed by many others]; therefore we should also expect the
egg production to be proportionately larger. Despite this, in the 5
years since protection, there has been no statistically significant increase
in recruitment levels on areas of the continental shelf adjacent to the
protected area (D.Hart, personal communication). Therefore, while there
is good evidence that reserves enhance egg production and that recruitment
should be enhanced, it will be difficult to actually show that this translates
into improved recruitment.
[don't even try]
In the case of the Georges Banks, longer time-series of data may be required.
Elsewhere much larger areas of marine reserves will be needed to achieve
increases of larval abundance on a scale that could be expected to show
measurable results.
[if it don't work, make it bigger!]
Take good notice, marine reserve skeptics
because for many years we have been bombarded with the opposing fallacy.
Another myth laid to rest?
The current proportion of marine reserves along the New Zealand coastline
is less than 0.1%, just a drop in the ocean when it comes to influencing
recruitment. While no evidence for such increases currently exists from
marine reserves, stock-recruitment relationships have been shown in some
invertebrate populations based on experimental manipulations at spatial
scales (~1km) similar to most marine reserves (Prince et al. 1988). Larval
export is an important issue that requires more satisfactory resolution
both for conservation and the management of fisheries. In this regard,
a consistent application of ecological assumptions is desirable.
Ecological principles explain why no
benefit from larval export can be expected. If you want more fish, then
the only methods that works is fishery management. Nothing else will.
While fisheries scientists are correct to point out the weakness of stock
recruitment relationships, the models of fish population dynamics on which
most of their management strategies are based depend either implicitly
or explicitly on the existence of the stock-recruitment relationship (Jennings
et al. 2001). [oops, false] New Zealand s
Fisheries Act (1996) requires fisheries to be managed so that they achieve
Biomass at Maximum Sustainable Yield or Bmsy, a concept that is explicitly
founded on the stock-recruitment relationship [oops,
false]. It is inconsistent to deny the potential usefulness of reserves
as a source of recruits while simultaneously basing traditional fisheries
management decisions on models that rely on the stock-recruitment relationship.
The author may be a little out of his
tree here. Only two paragraphs above, he explains that there is no such
stock-recruitment relationship, with which ecological principles agree
(for broadcast spawners). Fisheries models do not relate stock size
to recruitment.
The idea that marine reserves may provide insurance against recruitment
overfishing in the wider stock has been used (retrospectively) as the basis
for justifying the proportion of coastline that should be protected. Bohnsack
et al. (2003) has suggested that proportions of between 10-20% protection
[oops,
Bohnsack claims 20-30%] are prudent because protection of smaller
proportions of the coast requires unrealistically large compensatory increases
in larval survival [oops] under overfishing
conditions in unprotected areas (Fig.8).
The Bohnsack et al 2003 article makes
for amusing reading, underlining precisely the woolly thinking now favoured
by marine reserves scientists. Read Marine reserve
target sizes to get a grip on this and also to see how much disagreement
exists.
Reserves as management tools: insuring against the unexpected The (Fisheries) act has got all the tools to manage them (fisheries)
in a much more sophisticated and targeted way than the Marine Reserves
Act, which is a totally blunt instrument (N. Gibbs The Dominion, 7
February 2002)
The fatal flaw is the experience of the QMS (quota management
system). Over the past 17 years weve trashed a whole range of fish stocks
(B.
Weeber The Dominion, 7 February 2002)
New Zealand was one of the first countries to implement an output- or
quota-based fisheries management system for all major commercial species.
In this approach the amount of fish that can be caught sustainably in any
area is determined and then used as the basis for setting the quota. The
Quota Management System has been praised by analysts, both within New Zealand
and overseas, as a model of how fisheries should be managed since one of
its major results has been an end to the race for fish (Dewees 1998).
There is no doubt that the QMS is an improvement on previous systems that
tried to control harvest levels by limiting the number of fishers, types
of gear or fishing seasons. It is argued by some that with a cap on the
total amount of fish that can be taken, there is no need for marine reserves
as a means of managing fish populations. Setting aside concerns relating
to habitat destruction and indirect effects of fishing, there is some validity
to this line of argument. However, it hinges on the crucial assumption
that the quota level is in fact set at the right level.
As has been the case with all other attempts to manage fisheries, it
is becoming apparent that we do not always have perfect knowledge of fish
stocks that will allow us to set quotas correctly. Even with New Zealands
QMS, some stocks in some areas have been declining through the 1990s e.g.
Stewart Island and Catlins Paua Haliotis iris (Andrew et al. 2002),
Fiordland J.edwardsii (Starr et al. 1997), and Blue Cod in the northern
South Island (Ministry of Fisheries 2003). Even for the P.auratus fishery
in northeastern New Zealand (SNA1), major uncertainties remain surrounding
total catch estimates (Boyd and Riley 2002). The P.auratus fishery
in SNA1 is one of the most commercially and recreationally important inshore
fisheries in New Zealand. It is also one of the best studied, yet despite
this it has recently become apparent that the total P.auratus catch
exceeds the total allowable catch (TAC) by 40% or over 3000 tons (Table
2). This uncertainty has major implications for the setting of TAC.
Although by no means perfect, the QMS
does stop the race for fish while providing overarching control. An important
element is feedback from stock assessments and catch per unit effort
to set catch limits. There is room for more precautionary stock levels
as more experience is gained. Each of the problems the author may raise,
has its own explanation but overall, stocks are better managed than before.
An enormous amount of open discussion precedes each Total Allowable Catch
setting. In the last sentence, the author mentions the uncertain part the
recreational fishery plays. Recent assessments (although disputed) raised
the estimated recreational take by an unprecedented amount. Yet fish stocks
are not worse off.
The author does not realise that some of the
overfishing happened on stocks not yet in the QMS, for the purpose of establishing
a high catch history on the basis of which the first quotas are handed
out (for free). It is called 'catching quotas'.
Economic, demographic and technological trends indicate that the level
of recreational fishing is likely to continue to grow. Given the difficulty
of controlling and monitoring recreational fishing, marine reserves are
likely to provide an important level of insurance against this uncertainty.
They also provide a significant resource for those recreational users of
the marine environment who wish to enjoy their fish alive, and in their
natural habitat. Tourism based on the marine reserve at Leigh alone
is estimated to bring 100,000 visitors per year, contributing substantially
to the local economy (Cocklin et al.1998).
The insurance argument is a well established
fallacy giving peace of mind only to eco-activists, but what does it mean?
The idea is that even if all else fails (goes wrong) outside, the marine
reserve would still be there to seed its species in the bared areas outside.
Let's check this out.
When we think about insurance, we understand
that we pay a little (premium) in order to expect a large payout (indemnity)
when things go wrong. But marine reserves do not protect against large
scale events such as damage from hurricanes, invasions of introduced species,
oil spills, mass mortalities from poisonous plankton blooms, degradation
by mud and sewage and so on, because these play havoc equally inside and
outside marine reserves. What about global climate change, global warming,
ultraviolet irradiation, industrial pollution and acid rain? So who wants
to take out such an insurance?
Since no-take marine reserves affect only fishing,
they may provide an insurance for fishing-related accidents. But this only
relates to fished species that are resident, which is but a very small
part of the total. Who wants to take out such insurance?
Suppose a marine reserve does indeed provide
the larvae to restock the outside in case of a fishing related disaster,
it would take a very long period before stocks have re-established themselves,
often more than ten years. During all that time, the fishery has to be
discontinued. Who wants to take out such insurance, especially since better
methods are readily available? Is there anybody who believes that a protected
sea mount hundreds of sea miles from a damaged one will repopulate it?
Obviously, the insurance argument is seriously flawed.
Marine reserves skepticists, here is another
myth laid to rest. For many years we have been bombarded by the claim that
the Goat Island marine reserve attracts 250,000 visitors per year (DoC
repeatedly claims 300,000!). Now it is down to 100,000, and this already
in 1998! We claim that visitors stayed away in droves since DoC banned
the feeding of the fish (2000). Is that the cause of this sudden decline
or is there more readjustment to come? When will the myth of $12M/yr or
$120 per person per visit to the local economy bite the dust?
Fig.8.
Recruitment and minimum levels of reserve protection. The logistic population
growth curve a) in which fastest growth is achieved when the population
is at about half of its maximum level (B max ) is the basis for the principle
of maximum sustainable yield (MSY).
At higher levels of biomass population growth slows due
to density dependent effects on the survivorship or growth of recruits.
The New Zealand Fisheries Act 1996 requires fisheries to be managed so
that they are at the BMSY level b), forming the basis of setting TAC in
all quota management species. In a population harvested down to Bmsy, approximately
50% of the original spawning potential (egg production) of the population
would remain, but reduced egg production is expected to be compensated
for by a two-fold increase in egg survival due to release from density
dependent effects c).
Reducing the population to smaller and smaller fractions
of its spawning potential, requires a corresponding increase in egg survival
in order to maintain the original spawning potential and sustainable harvest.
At levels at or below 20% of spawning potential the increases in egg survival
required begin to exceed those that might be realistically expected (i.e.
5 10 fold increases). It has been argued by Bohnsack (2003) that protecting
at least 20% of a species habitat in reserves will provide insurance that
at least that much of the spawning potential remains.
[sigh] Armchair ecology on the
computer screen. Refer to the economies of exploitation in Resource
Management for a better explanation.
Table 2. Snapper fishery in northeastern New Zealand
(SNA1). Data (tons) are taken from Gilbert et al. (2000) except for estimates
of recreational catch in 2000 (Boyd and Riley 2003). TAC= total allowable
catch, TACC= total allowable commercial catch. *estimated based on most
recent previous estimate. not available at the time of publication.
TACC
Commercial Landings
Recreational Catch
TAC
Total Landings
1996
4,938
5,049
2,322
7,371
1997
4,500
4,519
2,322*
7,550
6,841*
1999
4,500
4,411
2,322*
7,550
6,733*
2000
4,500
6,200
7,550
10,700
Conclusion: Reserves as tools for science and management Fisheries management is broader than fisheries yield enhancement, or
even maintenance. This is a distinction that those debating marine reserves
frequently fail to make. Increasingly, modern fisheries legislation, including
the New Zealand Fisheries Act (1996) requires that not only fish stocks,
but also essential fish habitat, be managed sustainably. In order to achieve
this, complex interactions must be accounted for e.g. habitat protection,
by-catch species and changes in population structure. Given the difficulty
of modeling even one stock accurately, marine reserves are probably the
best way to achieve the goals of conservation, ecosystem management and
verifiably sustainable fisheries. Given the less-than-perfect track record
of even our best fisheries management systems, marine reserves are an option
that must [?]be implemented.
The author has just explained why marine
reserves do not help fisheries management, backed by scientific evidence.
Now he bases the need for marine reserves on the less than perfect track
record of fisheries management, losing out of sight the very disappointing
track record of coastal marine reserves. Due to coastal pollution, such
reserves can no longer protect the environment. We wonder how marine reserves
can reduce by-catch, since these arise from fishing. We also wonder how
they protect against seabird by-catches.
As far as population structure is concerned,
marine reserves may offer protection and restoration of old wise fish,
but this is not borne out in practice. For instance, spotted black grouper
and other serranids have not appeared in protected areas. Something else
is amiss.
Evidence from New Zealand suggests that their impacts on fishers and fishery
economics are at worst neutral [oops], but
that they also bring many benefits in terms of protecting habitat and populations
of exploited species [oops]. Reserves must
be used in combination with other management systems to achieve overall
protection of marine resources. For example, reserves may displace fishing
effort, which may have undesired effects on fished areas especially where
fishing overcapacity exists. Even though the level of impact is likely
to be minor if less than 20% of habitat is protected, population growth
and increasing access to marine environments mean that, even with reserves,
pressure on fish stocks and the marine environment will increase. Without
a range of other management tools (e.g. QMS) reserves may introduce
a false sense of security and exacerbate problems elsewhere. As with
anything new, there is a certain level of fear and ignorance surrounding
marine reserves. With time, it is likely to become more and more apparent
that we have nothing to lose and everything to gain from including marine
reserves as one of a range of essential management tools for the marine
environment. [sigh]
This whole article has failed to substantiate
why marine reserves are necessary. It has also failed to look at alternatives,
and it has failed to identify all matters wrong
with the present situation and it has failed to identify the main threats
to our seas. Ignorance, amazingly, reigns particularly with those who call
themselves scientists but who fail to observe the real threats to our seas
while being too arrogant to listen to what seamen have to say. Read FAQs
to avoid making the same mistakes. These scientists have succumbed to an
ideological course for political gains, rather than following the scientific
trail of discovery of how things work. In doing so they have been blind
to the obvious. Not willing to bite the hand that feeds them, they have
played the hand of a rogue government department which deems its advocacy
role more important than telling the truth. As a result, a whole generation
of people may lose their trust in Governance and science.
.
References.
In an article like this, which is obviously
aimed at the general public, one would expect references to be freely available.
However, this is not so. Scientists have created themselves an exclusive
ivory tower to which layman's access is forbidden. None of the articles
below are freely available. To make matters worse, the Department of Conservation
refuses access to scientific reports paid for by the NZ public purse. Under
threat of legal action and invocation of the Official Information Act,
we are gradually gaining access to these reports. So much for the Department's
legal obligation to engage in widespread consultation!
Department of Conservation scientific reports:
consult the Seafriends Library and
recent
additions.
Ministry of Fisheries: many reports on their
web site free of charge
University of Auckland: dissertations can be
read in the library but not copied.
Ecological Applications: with some difficulty
old editions can be accessed free of charge on the Web.
Marine & Freshwater Research: the RSNZ does
not provide these free of charge to non-members.
Adams, S., Mapstone,B.D., Russ,G.R.and Davies,C.R.2000. Geographic variation
in the sex ratio,sex specific size,and age structure of Plectropomus
leopardus (Serranidae)between reefs open and closed to fishing on the
Great Barrier Reef. Canadian Journal of Fisheries and Aquatic Sciences.
57:1448-1458.
Allison,G.W., Lubchenco,J. and Carr,M.H. 1998. Marine reserves are necessary
but not sufficient for marine conservation. Ecological Applications
8 (Suppl.):79-92.
Andrew,N.L. and Choat,J.H. 1982. The influence of predation and conspecific
adults on the abundance of juvenile Evechinus chloroticus (Echinoidea:Echinodermata).
Oecologia 54:80-87.
Andrew,N.L. and MacDiarmid, A.B.1991. Interrelations between sea urchins
and spiny lobsters in northeastern New Zealand. Marine Ecology Progress
Series 70:211-222
Andrew,N.L. and O Neill,A.L. 2000. Large-scale patterns in habitat
structure on subtidal rocky reefs in New South Wales. Marine and Freshwater
Research .51:255-263 .
Andrew,N.L., Naylor,J.R. and Kim,S.W. 2002 . Fishery independent surveys
of the relative abundance and size structure of paua (Haliotis iris
) in Pau 5B and PAU 5D. New Zealand Fisheries Assessment Report
2002/41.24 p. Ministry of Fisheries, Wellington.
Ayling,A.M.1978. Okakari Point to Cape Rodney Marine Reserve Survey.
98 p. Leigh Marine Laboratory Bulletin 1. University of Auckland, Leigh.
Babcock,R.C., Kelly,S., Shears,N.T., Walker,J.W.and Willis,T.J. 1999. Large-scale
habitat change in a temperate marine reserve. Marine Ecology Progress
Series 189:125-134.
Bentley,N., McNeill, S.E., Davies,N.M.and Davies C.R. 2001. An example
of assessing spatial closures as a fisheries management tool.12 p.
Draft Report to the New Zealand Ministry of Fisheries.
Bohnsack, J.A. 1997. Consensus development and the use of marine reserves
in the Florida Keys, USA. p.1927-1930. 8th International Coral Reef
Symposium, Panama. edited by H.A. Lessios and I.G.Macintyre. Vol2.
Bohnsack,J.A., Causey,B., Crosby,M.P., Griffis,R.B., Hixon,M.A., Hourigan,T.F.,
Koltes,K.H., Maragos,J.E., Simons,A. and Tilmant,J.T. 2003. A rationale
for minimum 20-30% no take protection. 9th International Coral Reef
symposium, Bali.edited by Kasim Moosa, M.K, et al. Bali Indonesia. in press.
Boyd,R.O. and Reilly,J.L. 2002. 1999/2000 national marine recreational
fishing survey: harvest estimates. Draft New Zealand Fisheries Assessment
Report. 28 p. Ministry of Fisheries, Wellington.
Choat, J.H and Schiel,D.R. 1982. Patterns of distribution and abundance
of large brown algae and invertebrate herbivores in subtidal regions of
northern New Zealand. Journal of Experimental Marine Biology and Ecology.
60:129-162.
Cocklin,C., Craw,M.and McAuley,I. 1998. Marine reserves in New Zealand:
use rights, public attitudes and social impacts. Coastal Management
26:213-231.
Cole,R.G. 1994. Abundance, size structure, and diver- oriented behaviour
of three large benthic carnivorous fishes in a marine reserve in northeastern
New Zealand. Biological Conservation 70:93-99.
Cole,R.G. and Keuskamp,D. 1998. Indirect effects of protection from
exploitation: patterns from populations of Evechinus chloroticus
(Echinoidea) in northeastern New Zealand. Marine Ecology Progress Series.173:215-226
Cole,R.G., Villouta E. and Davidson,R.J. 2000. Direct evidence of limited
dispersal of the reef fish Parapercis colias (Pinguipedidae) within
a marine reserve and adjacent fished areas. Aquatic Conservation: Marine
and Freshwater Ecosystems. 10:421-436.
Cole,R.G., Syms,C., Alcock,N.K., Notman,P., Stewart, R.and Carr,M.H. Sampling
abundance and size of temperate reef fishes in marine reserves: a methodological
comparison of open circuit scuba, semi-closed rebreathers and baited video.
in press.
Collie,J.S., Escanero,G.A. and Valentine,P.C. 1997. Effects of bottom
fishing on benthic megafauna of Georges Bank. Marine Ecology Progress
Series.155:159-172.
Crossland,J. 1976. Snapper tagging in North-East New Zealand, 1974:
Analysis of methods, return rates, and movements. New Zealand Journal
of Marine and Freshwater Research. 10:675-686.
Davidson,R.J., Villouta,E. Cole,R.G.and Barrier,R.G.F. 2002. Effects
of marine reserve protection on spiny lobster (Jasus edwardsii) abundance
and size at Tonga Island marine reserve, New Zealand. Aquatic Conservation:
Marine and Freshwater Ecosystems.12:213-227.
Dayton,P.K., Tegner,M.J., Edwards,P.B. and Kristin L.Riser. 1998. Sliding
baselines, ghosts, and reduced expectations in kelp forest communities.
Ecological Applications 8:309-322.
Denny,C., Babcock,R. 2002. Fish survey of the Mimiwhangata Marine Park,
Northland. Report to the Department of Conservation .28 pp.
Denny,C., Willis,T.J.and Babcock,R.C. 2002. Effects of Poor Knights
Islands marine reserve on demersal fish populations. 57p. Report to
the Department of Conservation.
Department of Conservation, 2002. Marine Reserves Bill Has First Reading.
http://www.doc.govt.nz/whats-new/presult.asp?prID=884
Dewees,C.M. 1998. Effects of individual quota systems on New Zealand
and British Columbia fisheries. Ecological Applications. 8 (Suppl.):133-138.
Edgar,G.J. and Barrett,N.S. 1997. Short term monitoring of biotic change
in Tasmanian marine reserves. Journal of Experimental Marine Biology
and Ecology 213:261-279.
Edgar,G.J. and Barrett,N.S. 1999. Effects of the declaration of marine
reserves on Tasmanian reef fishes, invertebrates and plants. Journal
of Experimental Marine Biology and Ecology 242:107-144.
Egli,D.P., and Babcock R.C. 2002. Optimising marine reserve design in
New Zealand Part I: Behavioural data for individual based models.
39 p. Report to the Department of Conservation, Northland Conservancy
Fowler Walker,M. and Connell,S.D. 2002. Opposing states of subtidal
habitat across temperate Australia: consistency and predictability in kelp
canopy-benthic associations. Marine Ecology Progress Series 240:49-56
Francis,M.P. 1993. Does water temperature determine year class strength
in New Zealand snapper (Pagrus auratus, Sparidae)? Fisheries
Oceanography 2:65-72.:
Francour,P., Harmelin,J.-G., Pollard,D., Sartoretto,S., 2001. A review
of marine protected areas in the northwestern Mediterranean region: siting,
usage, zonation and management. Aquatic Conservation: Marine and Freshwater
Ecosystems 11:155-188.
Freeman,D.J. and Duffy,C.A.J. 2003. Te Angiangi marine reserve: reef
fish and rock lobster monitoring 1995-2001. Department of Conservation,
East Coast Hawke s Bay Conservancy.
Gilbert, D.J., McKenzie,J.R., Davies,N.M. and Field,K.D. 2000. Assessment
of
the SNA 1 stocks for the 1999-2000 fishing year. New Zealand Fisheries
Assessment Report 2002/38. 52 p. Ministry of Fisheries, Wellington.
Green Party, 2002. Greens want more marine reserves. http://onenews.nzoom.com/onenews_detail/0,1227,100795-1-8,00.html
Harley,S.J. and Gilbert,D.J. 2000. Assessment of the Tasman and Golden
Bays snapper fishery for the 1999-2000 fishing year. New Zealand Fisheries
Assessment Report 2002/28. 42 p. Ministry of Fisheries, Wellington.
Hillborn,R. 2002. Marine reserves and fishery management. Science
295:1233-1234.
Jennings,S., Kaiser,M.J. and Reynolds,J.D. 2001. Marine Fisheries Ecology.
417 p. Blackwell Science Oxford
Jones,G.P., Cole,R.C. and Battershill,C.N. 1992. Marine reserves: do
they work? pp.29-45. in Proceedings of the Second International Temperate
Reef Symposium edited by C.N.Battershill et al. NIWA, Wellington.
Jones,G.P., Milicich,M.J. Emslie,M.J. and Lunow,C. 1999. Self-recruitment
in a coral reef fish population. Nature 40:802-804.
Kelly,S.1999. 1999 Lobster survey of Te Awaatu (The Gut) marine reserve,
Doubtful Sound. 12 p. Report to the Department of Conservation.
Kelly,S. 2000. 2000 Lobster survey of the Cathedral Cove (Te Whanganui
A Hei) marine reserve. 15 p. Report to the Department of Conservation.
Kelly,S., 2001. Temporal variation in the movement of the spiny lobster
Jasus
edwardsii. Marine and Freshwater Research 52:323-331.
Kelly,S., MacDiarmid,A.B., and Babcock,R.C., 1999. Characteristics of
spiny lobster, Jasus edwardsii, aggregations in exposed reef and
sandy areas. Marine and Freshwater Research 50 :409-416
Kelly,S., Scott,D., MacDiarmid,A.B. and Babcock,R.C., 2000. Spiny lobster,
Jasus
edwardsii, recovery in New Zealand marine reserves. Biological
Conservation 92:359-369.
Kelly,S. and Haggitt,T. 2002. Cape Rodney to Okakari Point marine reserve
lobster monitoring program: May 2002 Survey. 25 p. Report to the Department
of Conservation.
Kelly,S., Scott,D. and MacDiarmid,A.B. 2002. The value of a spillover
fishery for spiny lobsters around a marine reserve in northern New Zealand.
Coastal Management 30:153-166.
Kramer,D.L. and Chapman,M.R. 1999. Implications of fish home range size
and relocation for marine reserve function. Environmental Biology of
Fishes 55:65-79.
Lauck,T., Clark,C.W., Mangel,M. and Munro,G.R. 1998. Implementing the
precautionary principle in fisheries management through marine reserves.
Ecological Applications 8 (Suppl.):72-78.
Lough,P. 2002. Editorial Comment. Seafood New Zealand September:1-2
. Ministry of Fisheries. 2003. Blue Cod 7 Consultation. 24 p. http://www.mfish.govt.nz/current/blue-cod/index.html.
New Zealand Ministry of fisheries, Wellington.
MacDiarmid,A.B. and Breen,P.A.1982. Spiny lobster population change
in a marine reserve. pp.47-56 in Proceedings of the Second International
Temperate Reef Symposium edited by C.N.Battershill et al. NIWA, Wellington.
Murawski,S.A., Brown,R. Lai,H.L. Rago,P.J.and Hendrikson,L. 2000. Large-scale
closed areas as a fishery management tool in temperate marine systems:
the Georges Bank experience. Bulletin of Marine Science. 66 :775-798.
New Zealand Fishing Council.2001.Opposition to marine reserve call.
http://onenews.nzoom.com/onenews_detail/0,1227,70663-1-7,00.html
O Dor,R.K., Aitken,J.P., Babcock,R.C., Bolden,S.K., Seino,S., Zeller,D.C.,
and Jackson,G.D. 2001. Using radio-acoustic positioning and telemetry
(RAPT) to define and assess marine protected areas (MPAs). in: Electronic
tagging and tracking in marine fisheries. Methods and Technologies
in Fish Biology and Fisheries,Vol 1, edited by J.Sibert and J. Nielsen.
Kluwer Academic Press, Dordrecht.
Option4. 2002. What does option4 stand for? http://option4.co.nz/whatdoes.htm
Parrish,R.1999. Marine reserves for fishery management: why not.
California Cooperative Oceanic and Fisheries Investigations 40 :77-86.
Parsons,D., Babcock,R., Willis,T. 2000. Space utilization characteristics
of snapper (Pagrus auratus) in a marine reserve. Report to the
Department of Conservation. 31 pp.
Pauly,D., Christensen,V., Dalsgaard,J., Froese,R. and Torres,F.Jr. 1998.
Fishing
down marine food webs. Science 279:60-863.
Prince,J. 1995. Limited effects of the sea urchin Echinometra matthaei
(de Blainville) on the recruitment of benthic algae and macroinvertebrates
into intertidal rock platforms at Rottnest Island, Western Australia.
Journal of Experimental Marine Biology and Ecology.186:237-258.
Prince,J.D., Sellers,T.L., Ford,W.B. and Talbot S.R.1988. Confirmation
of a relationship between the localized abundance of breeding stock and
recruitment for Haliotis rubra (Mollusca: Gastropoda). Journal
of Experimental Marine Biology and Ecology. 1988. 122:17-24.
Roberts,C.M. 1997. Ecological advice for the global fisheries crisis.
Trends in Ecology and Evolution 12:35-38. Roberts,C.H. and Hawkins,J.P.
2000. Fully protected marine reserves: a guide .131p.WWF Endangered
Seas Campaign, Washington DC
Roberts,C.M., Bohnsack,J.A., Gell,F. and Goodridge,R. 2001. Effects
of marine reserves on adjacent fisheries. Science 294:1920-1923.
Roughgarden,J.1998. How to manage fisheries. Ecological Applications.
8:160-164.
Russ,G.R. and Alcala,A.C. 1996. Do marine reserves export adult fish
biomass? Evidence from Apo Island, central Philippines. Marine Ecology
Progress Series 132 :1-9.
Sala,E., Aburto-Oropeza,O., Paredes,G., Parra,I., Barrera, J.C. and Dayton,P.K.
2002. A General Model for designing networks of marine reserves.
Science 298:1991-1993.
Schmidt,K.F.1997. No-take zones spark fisheries debate. Science,
277 :489-491.
Shears,N.T. and Babcock,R.C. 2002. Marine reserves demonstrate top-down
control of community structure on temperate reefs. Oecologia.132 :131-142.
Shears,N.T. and Babcock,R.C. 2003. Continuing trophic cascade effects
after 25 years of no-take marine reserve protection. Marine Ecology
Progress Series 246:1-16.
Sladek Nowlis,J. and Roberts,C.M. 1999. Fisheries benefits and optimal
design of marine reserves. Fishery Bulletin 97:604-616.
Starr,P.J., Breen,P.A., Hilborn,R.H. and Kendrick,T.H. 1997. Evaluation
of a management decision rule for a New Zealand rock lobster substock.
Marine and Freshwater Research. 48:1093-1101.
Steinberg,P.D., Estes,J.A. and Winter,F.C. 1995. Evolutionary consequences
of food chain length in kelp forest communities. Proceedings of the
National Academy of Science USA. 92:8145-8148
Te Ohu Kai Moana. 2001. Marine reserves need to be put on hold.
http://www.tokm.co.press/10-12-01_Marine_reserves_ need_to_be_put_on_hold.htm
Thrush,S.F. Hewitt,J.E. Cummings,V.J. Dayton,P.K 1995 . The impact of
habitat disturbance by scallop dredging on marine benthic communities:
What can be predicted from the results of experiments. Marine Ecology
Progress Series.129:141-150 . Thrush,S.F., Hewitt,J.E., Cummings,V.J.,
Dayton,P.K., Cryer,M., Turner,S.J., Funnell,G.A., Budd,R.G., Milburn, C.J.
and Wilkinson,M.R 1998. Disturbance of the marine benthic habitat by
commercial fishing: Impacts at the scale of the fishery. Ecological
Applications. 8:866-879.
Turpie,J.K., Beckley,L.E. and Katua,S.M. 2000. Biogeography and the
selection of priority areas for conservation of South African coastal fishes.
Biological Conservation. 92:59-72.
Watling,L. and Norse,E.A.1998. Disturbance of the seabed by mobile fishing
gear: A comparison to forest clearing. Conservation Biology 12:1180-1197.
Willis,T.J. and Babcock,R.C. 2000. A baited underwater video system
for the determination of relative density of carnivorous reef fish.
Marine and Freshwater Research 51:755-763.
Willis,T.J., Millar,R.B. and Babcock,R.C. 2000. Detection of spatial
variability in relative density of fishes: comparison of visual census,
angling, and baited underwater video. Marine Ecology Progress Series
198:249-260.
Willis,T.J., Parsons,D.M. and Babcock,R.C. 2001. Evidence for site fidelity
of snapper (Pagrus auratus) within a marine reserve. New Zealand
Journal of Marine and Freshwater Research 35:581-590.
Willis,T.J., Millar,R.B. and Babcock,R.C. 2003. in press. Protection
of exploited fishes in temperate regions: high density and biomass of snapper
Pagrus
auratus (Sparidae) in northern New Zealand marine reserves. Journal
of Applied Ecology 40:214-227.
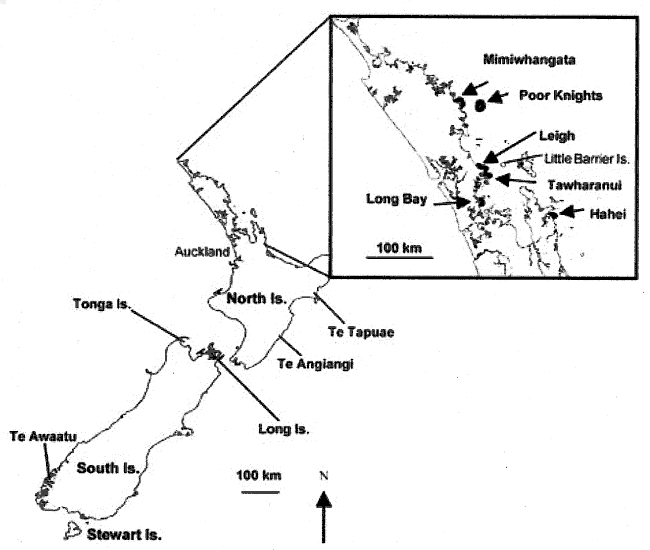
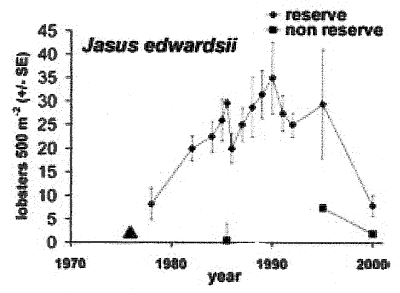 Fig.2.
Recovery of spiny lobster population in the Leigh marine reserve. While
no data were collected prior to protection and were only sporadic until
the mid 1990s the increase in density is most likely due to protection
and has been observed in other reserves around the North Island. The reason
for a substantial decrease in numbers at the Leigh reserve since 1995 is
not known but has not been observed at other reserves (e.g. Hahei). Data:
Ayling 1978, McDiarmid and Breen 1992, Kelly et al. 1996, Kelly and Haggitt
2000. Triangle= time of reserve creation (1976).
Fig.2.
Recovery of spiny lobster population in the Leigh marine reserve. While
no data were collected prior to protection and were only sporadic until
the mid 1990s the increase in density is most likely due to protection
and has been observed in other reserves around the North Island. The reason
for a substantial decrease in numbers at the Leigh reserve since 1995 is
not known but has not been observed at other reserves (e.g. Hahei). Data:
Ayling 1978, McDiarmid and Breen 1992, Kelly et al. 1996, Kelly and Haggitt
2000. Triangle= time of reserve creation (1976).
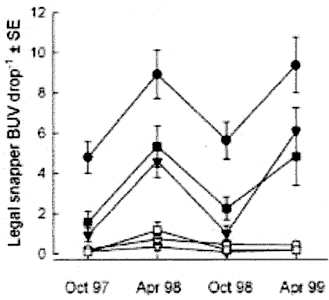
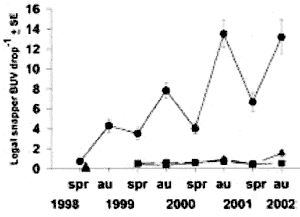
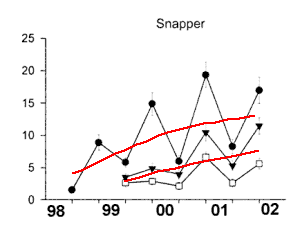
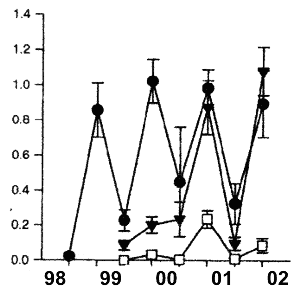
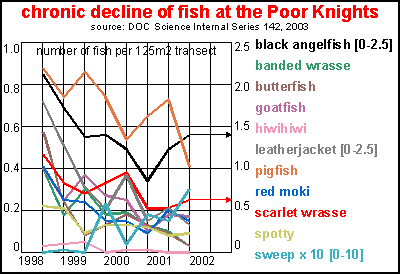
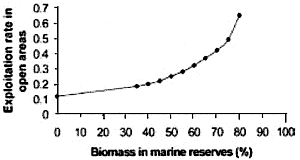

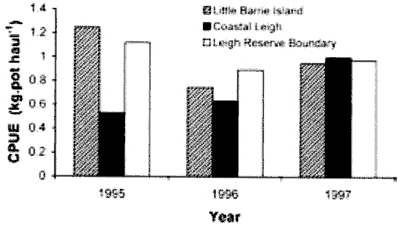
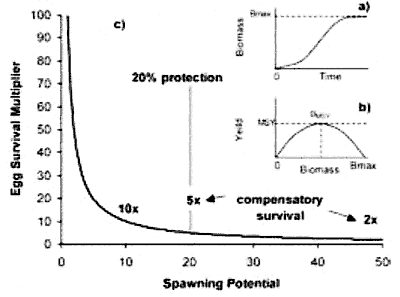 Fig.8.
Recruitment and minimum levels of reserve protection. The logistic population
growth curve a) in which fastest growth is achieved when the population
is at about half of its maximum level (B max ) is the basis for the principle
of maximum sustainable yield (MSY).
Fig.8.
Recruitment and minimum levels of reserve protection. The logistic population
growth curve a) in which fastest growth is achieved when the population
is at about half of its maximum level (B max ) is the basis for the principle
of maximum sustainable yield (MSY).